目 次
第1章地球形成の秘密
#原始地球の形成 #マグマオーシャン #地球の公転と自転の秘密
#地軸傾きの秘密 #地球の大きさと質量、密度 #地球の構造
第2章地球大気と海の秘密
#大気の形成 #海の形成の秘密 #太陽と地球の位置の秘密 #大気に酸素生成
#オゾン層の形成 #地球磁場の成立 #太陽風と磁気バリア
第3章大陸生成と邸陸移動の秘密
#最初の陸地 #始生代の地質 #大陸生成 #原生代初期の地表
#超大陸の形成― ロディニア大陸 #プレートテクトニクス
#プルームテクトニクス #スーパーホットプルーム
第4章全地球凍結の秘密
#スノーボールアース #スノーボールアースの推移 #寒冷気候と温暖気候の循環
#氷河期 #過去の大きな氷河期 #氷河期の原因 #海水準の変化
第5章生命誕生の秘密
#生命の定義 #極限状況での生命誕生 #極限環境に生きる古細菌
#人と共生する真正細菌 #全ての生物 ―真核生物 #多細胞生物ま誕生
#植物の多細胞体制 #動物の多細胞体制 #カンブリア爆発 #カンブリア爆発の原因
第6章 生命絶滅の秘密
#ビッグファイブ ―生物大量絶滅 #オルドビス紀末の大量絶滅
#デボン紀後期(F-F境界)の大量絶滅 #ペルム紀末(P-T境界)の大量絶滅
#陸上の動物と植物の絶滅 #超酸素欠乏- スーパーアノキシア
#三畳紀末(T-J境界)の大量絶滅 #白亜紀末(K-Pg境界)の大絶滅
第7章生命進化の秘密
#大型魚類の誕生 #大森林の誕生 #巨大昆虫 #爬虫類
#爬虫類の多様化 #恐 竜 #巨大な恐竜の誕生 #両生類の誕生
#大型草食性恐竜 #最大級の肉食恐竜 #哺乳類の時代 #始祖象の誕生
#ウマ科動物の誕生 #第三紀の動物 #種の形成 #進化論
#チャールズ・ダーウィンの進化論 #ホミニンの誕生と拡散
原始地球の形成
地球は約46億年前、他の太陽系の惑星と同様、太陽の誕生とともに、その周囲にできた原始惑星系円盤の中で形成された。
太陽系が形成される少し前に、近くで超新星爆発があり、爆発の衝撃が引き金となって、星間物質の収縮が始まり、太陽系の形成が始まった。
力学的シミュレーションで、原始太陽系がガスや塵の状態から多数の微惑星(サイズは数㎞以上)を経て、惑星サイズまで成長するのに数百万年から数千万年かかった。
隕石の多くは、この時に生まれた微惑星のかけら(始原的隕石)である。太陽系形成が始まって10万年~100万年で、現在の地球の軌道周辺には微惑星が衝突・合体して形成された数十個の「月から火星サイズの惑星胚」が生じ、各々の軌道を廻るようになる。

原始惑星系円盤のほとんどは、水素やヘリウムなどのガスからできていたが、わずかながら宇宙塵を含んでいた。原始地球のサイズが、火星レベル、現在の地球質量の10分の1になると、衝突のエネルギーで微惑星は融解する。
数十個の惑星胚は、お互いの重力で軌道が乱れ、その結果軌道が交差して衝突を繰り返す。このレベルの衝突を、ジャイアントインパクトと呼ぶ。

地球形成時に、いくつか起こったジャイアントインパクトの最後の衝突で月ができた。
この時の衝突エネルギーは非常に大きく、衝突後の地球と月は、双方とも全体が溶融状態にあった可能性が高い。
放射性元素ハフニウム182に関する詳細な分析で、地球と月のマントルの形成が始原的隕石形成の約3千万年後であったと報告されている。
また地球全体が溶融したため、核を形成する鉄とマントルとなるケイ酸塩成分の分離と、鉄成分の地球中心部への沈降で、現在見の地球の層状構造が始まった。
なお地球の年齢は、地球岩石を、ウラン・鉛年代測定法では45億年から46億年、隕石をウラン・鉛年代測定法やルビシウム-ストロンチウム法では45~46億年という数値が出ている。 #目 次
マグマオーシャン
太陽から3番日の、岩石天体である初期地球は、その大きさが現在の半分くらいのころ、微惑星の残骸(隕石)との絶え間ない激しい衝突や、内部放射性物質の壊変、重力の凝縮効果による内部圧力の増加のために、極度な高熱であった。
微惑星の衝突エネルギーによる発熱によって、地球表面から半径の3~4割ほどの深さまで、地球は溶融状態であった。これをマグマオーシャン(マグマの海)と呼ぶ。
この猛烈な加熱の結果、中に閉じ込められていた鉄と二ッケルが融けはじめて、化学的分化という過程が起きた。

重力の影響で重金属の液体の塊は、地球の中心に向かって沈み、地質年代の規模としてはかなり素早く、高密度の鉄の塊になった。
これと同時に、地球の溶融、いわゆる「火の玉」地球の状態で、融けた岩塊の軽い部分が地表に向かって上昇して凝固しはじめ、地表で薄い層の原始地殻となった。
この地殻の岩石物質に含まれていたのは、相当量のケイ素とアルミニウム、そしてそれより少ない量のカルシウム、ナトリュウム、カリウム、マグネシウム、鉄、さらに金や鉛やウラン(ウラニウム)を含む重金属であった。
分化が起きた後、原始地殻は浸食によって消失したり、大きく変わったりしたため、元々の構造の直接証拠となるものはほとんど残っていない。
放射年代測定で、最初期の大陸地殻が形成された正確な時期については、地質学者が現在でも調査を行っている課題である。 しかしながら、これまでに見つかった最古の岩石は、放射年代測定でおよそ40億年前となっており、原始地球も遅くても40 億年前には形成されていたと推定されている。
マグマオーシャンからは気体が抜け出たが、その中には水(H2O)や塩化水素(HCl)など、さまざまなものを含んでいた。これが原始大気である。
冷却が進みマグマオーシャンの表層が固結し、さらに地表の温度が水の臨界温度64度K(飽和蒸気が液化する温度)以下になると、水蒸気が凝縮して原始海洋を形成した。そこでは塩化水素(塩酸
―高温の水にも溶ける)が溶解し、希塩酸の海になった。

しかし、ただちに海底の岩石と反応して中和され、塩化ナトリウム(NaCl)を主体とする海水ができた。地球ができてから8億年以内に、海はできていたと考えられている。その理由は、少なくとも38億年前には、海底にたまった堆積岩が有ったことが分かっているからである。
一般の岩石は、大部分が珪酸塩鉱物の集合体で、少量の酸化物・炭酸塩鉱物・硫化物などを含んでいる。 珪酸塩鉱物とは、珪酸(SiO2)とある金属酸化物が結びついてできた化合物である。#目 次
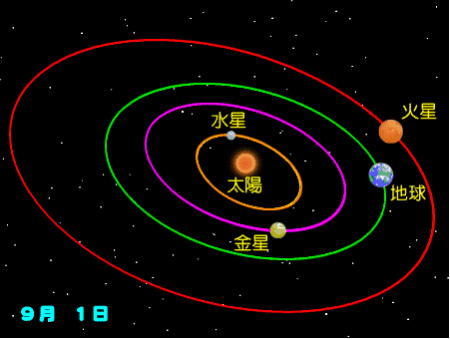
地球の公転と自転の秘密
・公転
地球の自転、公転ともに、天の北極から見て反時計回りである。
地球の自転は、最期のジャイアント・インパクトによって起きたと考えられる。
公転は、太陽の引力と太陽の自転の影響であり、互いの重力に引かれて共通重心の周りを公転している。当然、太陽も反時計回りで自転している。
円に近い楕円形の軌道を描き、太陽の周りを、1太陽年に1回公転し、また0.9973平均太陽日に1回自転している。
1太陽年とは、太陽が春分点から春分点まで、一巡りする時間、すなわち季節が一巡する時間をいい、約365.242 日である。
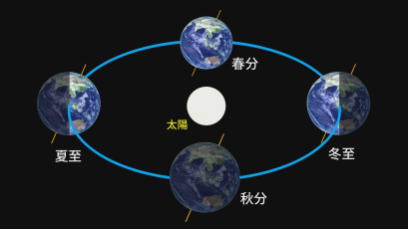
地球の歳差(春分点が恒星に対し、毎年50秒余ずつ西方へ移動する現象)により、春分点が移動するため、1太陽年は、恒星が動かないものとして見た時に、地球が太陽の周りを一周する時間として定義される1年(恒星年)より短い。
つまり1 恒星年は約365.256 日である。
太陽日とは、太陽が子午線を通過してから、翌日再び通過するまでの時間を真太陽日という。
この楕円の形は、10万年ほどの周期で、変化することが分かっている。楕円の軌道離心率(天体軌道を決めるパラメータで、どれだけ円から離れているかを表す値)は、0.0167である。
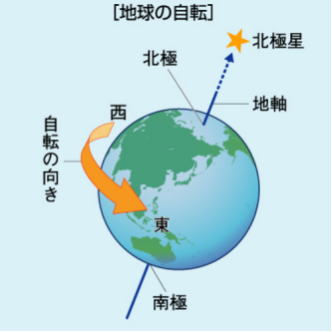
・自転
地球が自転して、元の位置に戻るまでの時間を「自転周期」といい、2種類ある。
一つは、恒星に対してのもので、これを恒星日という。
正確には、春分点が南中(太陽が真南にくる)してから、次に南中するまでの時間をいう。恒星日は、平均として23時間56分4秒である。
二つは、太陽に対してのもので、これを平均太陽日 (LOD) という。厳密には、天の赤道上を等速運動するとした仮想太陽(平均太陽)が、南中してから次に南中するまでの時間をいう。
日常生活では、平均太陽日の方が重要であり、時間の単位(国際単位)、又は暦の単位である「日」はこれに基づいている。
平均太陽日(LOD―日常の「1日」)の長さは、24時間ちょうどではなく、24時間 + 1から2㎜秒程度である。
LODの長さは一定ではなく、日々変動している。これに対して、時間の単位としての日は、正確に8万6千400秒である。
1太陽年や1恒星年を表現するのに用いられる1日は、太陽系天体の位置計算で、時刻引数として使用される力学時の1日で、1平均太陽日と考えても特に問題はない。
平均太陽日は、6億年前には約22時間であり、その時点での1年は、約400日だった。
地球誕生直後の1年は、2千日相当と考えられる。このように、かつて早かった自転速度は、徐々に遅くなっている。
これは、月や太陽の引力によって起こる、「潮汐作用」で動く海水が、自転運動よりも遅く、摩擦抵抗として作用すからである。
このため10億年後には、地球自転は31時間になると試算されている。
ただし、短期的(50年~ 100年程度)には、必ずしも長くなっているわけではなく、この30年間では短くなっている。 #目 次
地軸傾きの秘密
地軸とは、地球が自転する中心軸であり、北極点と南極点を結ぶ直線を指す。 地球以外の惑星及や衛星でも自転する中心軸を地軸と呼ぶ。
地球の地軸は、公転面の法線に対して、約23・43度傾いている。
この地軸の傾きは、原始地球が形成されるとき、何度かのジャイアント・インパクトと、最期のジャイアント・インパクトによって傾いた。
そのときの衝突エネルギーで飛び散ったガス状物質が、やがて重力で固まり月を形成した。そしてその形成された月の引力作用で、地球の回転軸が、傾いたまま安定した。この地軸の傾きで、地球では安定した季節変化が生じている。
惑星の季節変化の主な要因は、軌道離心率と自転軸の傾きが考えられるが、地球の場合は自転軸の傾きが大きい。
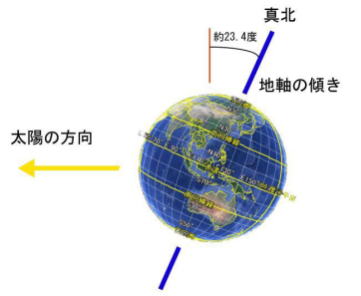
地球の軌道離心率が0.0167ということは、太陽に最も接近したとき(近日点通過)と、太陽から最も遠ざかったとき(遠日点通過)で、太陽約3個分の距離が違うことを意味している。(0.01天文単位が太陽直径程度である)
地球に対する太陽光量では、約7%が変動する。これよりも、自転軸の傾斜を原因とする太陽高度の変化(光が差し込む角度)と、日照時間が効くのである。
太陽に最も接近するのは、1月4日前後、最も離れるのは7月5日前後である。離心率や自転軸の傾斜は、木星などの引力の影響により、数万年周期で変動している。
地球の赤道の傾きは、22~24度の範囲を、およそ4万1千年の周期で変化している。
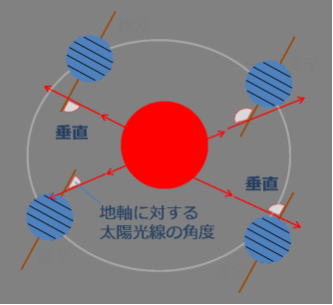
もしも地軸に傾きがなければ、それぞれの緯度で、季節の移り変わりがなくなり、緯度間の気候の差が顕著になり、地球全体の気候はもっと激しいものになっただろう。
一方で、傾きが強すぎれば、気候条件は混沌としたものになる。地球に生命の誕生に必要な、絶好の地軸の角度をもたらしたのは、月の影響である。
つまり地軸が傾いていることから、夏季には日が高く昇り、昼の時間が長く、冬季には日が低く、昼が短い。
単位面積当たりの太陽エネルギーの照射量と、日照時間とが変化することで、季節が生じる。
北緯23・43度(23度26分)を走る北回帰線上では、1年に1度、夏至に太陽の南中高度が90度になるが、これも地軸の傾きによる。
逆に90~23・43の北緯66・56度(66度33分)の北極圏では、夏至に太陽が沈まず(白夜)、冬至に日が昇らない。
なお、南半球では、暦と季節との対応が反対(6月は冬、12月は夏)となるため、南回帰線上では、12月の冬至(現地の夏)に太陽の南中高度が90度となり、南極圏では冬至を含む12月前後(現地の夏)に白夜となる。
地軸は、公転の影響を受けないため、常に一定の方向を指す。そのため、天球の北半球と、地軸の交点付近の恒星である北極星は、1日のどの時刻でも、1年のどの季節でも、同じ高度(角度)に見える。この高度は、観測者の居る地点の緯度によってだけ決まるので、北半球では、北極星の高度を測定すれば、地球上のどの地点でも緯度を計算できる。
このため、数百年にわたって、北極星が航海などに役立ってきたのは、こうした地軸の傾きによる。ただし、非常に長い期間を想定した場合、地軸自体の指す方向は変化する。これを歳差運動と呼ぶ。
歳差運動自体は珍しいものではなく、コマの首振り運動のように日常観察できるものである。地軸の歳差運動の周期は、約2万5千800年である。このため、北極星に該当する恒星も、相対的に変化しているように観察される。 #目 次
地球の大きさと質量、密度
・地球の大きさ
地球の形は、ほぼ回転楕円体で、赤道の半径は6千378㎞。
極半径は、6千357㎞。 地球の運動は、365日強で太陽の周囲を1周し、24時間で1回自転し、太陽からの平均距離は1億4千960万㎞である。
地球の内部は大まかに地殻、マントル、核の3部分から成っている。地球全体の平均密度は、1㎝立方当たり5・514gである。表面は大気に覆われている。
太陽系内の惑星では、2天文単位内に存在し、岩石質外層と鉄を主成分とする中心核を持つ「地球型惑星」に分類される。太陽系の地球型惑星の中での大きさ、質量ともに最大である。

地球の組成は、地表面からの深さによって異なる。
地殻に存在する元素は、酸素(質量比49.5%)とケイ素(同25.8%)が主体で、以下アルミニウム・鉄・カルシウム・ナトリウム・カリウム・マグネシウムなどの金属元素が含まれる。この元素別質量百分率は、クラーク数として纏められている。
ほとんどはケイ酸塩など、金属酸化物の形で存在する。
対照的に、中心部分は鉄やニッケルが主体である。
地表面の71・1%は液体の海で被われており、地表から上空約百㎞までの範囲には、窒素・酸素を主成分とする大気がある。大気の組成は高度によって変化する。
地球の質量と密度
地球の質量は5・972×1024 kgである。
万有引力定数と、地心重力定数から計算される。体積と質量から、平均密度が求められる。これは水の5・5倍、花崗岩の2倍、鉄の0・7倍程度に相当する。
地球は、太陽系で最も密度の高い惑星である。
逆に、一番密度が低いのは土星である。また水星や金星の密度は、地球に近い。
地球を構成する物質の種類と分布を探るには、地球内部での圧力上昇によって圧縮される程度を考慮して、1気圧下の密度に直す必要がある。このような補正を加えると、地球の平均密度は、約4千100立方㎏になる。
地球以外の惑星の内部構造は、観測データがなく、推定された補正平均密度は、水星は約5千400立方㎏、金星は地球とほぼ同じで約4千100立方㎏、火星は約3千800百立方㎏である。
これら補正された平均密度の違いは、金属の含有量の違いを反映している。一見、金属量は、太陽から離れるにしたがって減少するように見えるが、その理由はわかっていない。
#目 次
地球の構造
地球の層は、化学組成と物理的特性の二つによって定義することかできる。
地球は地殻、マントル、中心核(コア)からなる。さらに、地殻は海洋地殻と大陸地殻に分けられる。
薄い海洋地殻は、主に融解した物資が凝固してできた、黒い火成岩(玄武岩)からできている。その厚さは、およそ8㎞。これよりも厚い大陸地殻は、平均して40㎞の厚さがあり、山岳地方ではでは60㎞以上.にもなる。
大陸地殻には、多様な岩石がたくさん含まれているが、一般に上部は花崗岩、下部は玄武岩からなっている。
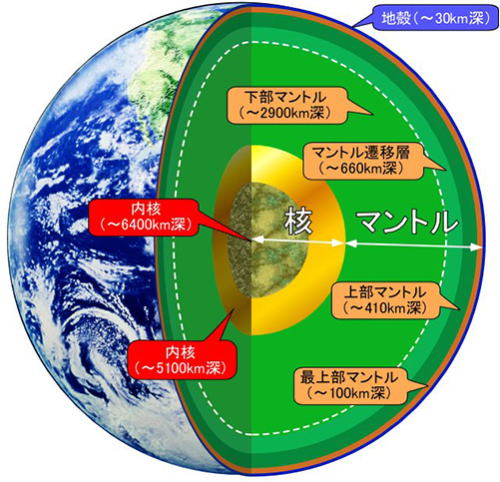
地球体積の80%以上を占めているのがマントルである。
深さ2千900㎞まで続く硬い岩石がある部分のことで、地殻とマントルか接する部分では、化学組成に大きな変化が見られる。部マントルは、マグネシウムに富んだ粗粒火成岩の橄(かん)欖(らん)岩(がん)からなっている。これは、かなり深いところで圧縮されて、高密度の結品構造になったもの。
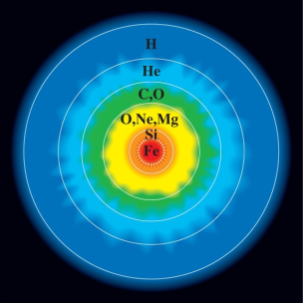
中心核(コア)の化学組成は、鉄とニッケルの合金と見られ、鉄と化合物を作るほかの元素も存在している。 コアの極度の圧カ下では、これらの元素の密度は、水の密度のおよそ14倍になっている。
地球の構造の物理的特性は、深さとともに増加する密度のほか、中心に向かうにつれて劇的に高まる温度の影響を受ける。
深さ100㎞の地球の温度は、約1千200~1千400℃で、中心核の温度は、6千700℃以上と推定されている。中心核が並外れて高温であることは、地球の形成時に、隕石の降着で得た熱エネルギーの大半を、保持してきたことを物語っている。
地球の主な層は、物理的特性によって5つに分けられる。
リソスフェア(地殻および上部マントルのごく一部)、アセノスフェア(マントルでより深くて熱い部分)、メソ.スフェア(アセノスフェアより下で、外核より上の部分)、そして外核および内核である。
リソスフェア(岩石圏)とアセノスフェア(岩流圄)は、地球の一番外側にある層で、地殻と上部マントルを形成しており、冷たくて非常に硬い部分として一体となって機能している。
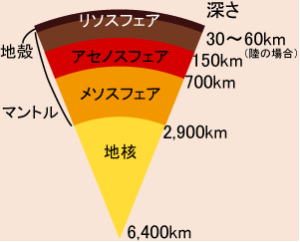
リソスフェアの厚さは、場所によって変わるが、だいたい100㎞内外であるのに対し、その下のアセスノフェアは、上部マントルを通って豹2200㎞、場所によっては700㎞の深さまである。
アセノスフェアの上部には、溶融が起きるかなり高温の場所があり、そのために上にあるリソスフェアは、アセノスフェアとは独立に動く。この事実は、プレートテクトニクスで、非常に重要な意味を持っている。メソスフェア(中問圏)は、下部マントルの層で、流体的なアセノスフェアの下にあって、高温であるにもかかわらず、圧力で押し固められている。こうしてアセノスフェアより硬いメソスフェアは、深さ約700㎞~2千900㎞の間に位置している。
そこから更に深い所にあるのが、外核と内核で、それぞれの力学的特性は大きく異なる。外核はおよそ2千300㎞の厚さがある液体の層である。外核内では、液体の金属鉄が対流していることにより、地球の磁場が生じている。半径約1千200㎞の球形の内核は、かなりの高温にさらされているが、相当な圧力がかかっているため、固体のような振る舞いをする。
#目 次
大気の形成
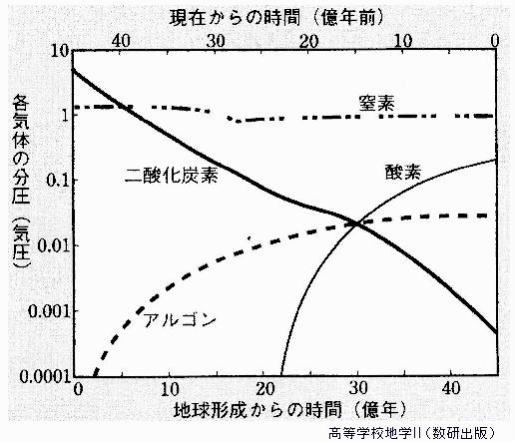
46億年前、誕生したばかりの地球の大気は、高温・高圧の水蒸気が大部分を占め、その他に二酸化炭素、窒素などを含んでいたと考えられる。 46億年前頃の原始大気は、主にヘリウムと水素からなり、高温高圧だった。これは、現在の太陽の大気と似た成分である。
また、水蒸気も含まれていて、その温室効果が、原始地球を高温高圧に保っていたという説もある。しかし、これらの軽い成分は、原始太陽の強力な太陽風で、数千万年のうちに、ほとんどが吹き飛ばされてしまった。

やがて太陽風は、太陽の成長とともに次第に弱くなってくる。
その後、数億年かけて地表が冷え、水蒸気が雨となって地表に降り注いで海ができる。すると大気の主成分は、二酸化炭素と窒素になり、さらに海に二酸化炭素が溶け込み、その一部がカルシウムイオンと結合して、石灰岩(炭酸カルシウム)として海底に堆積する。
こうして、大気中の二酸化炭素は減少し、大気の主成分は窒素になった。

この頃には、地表温度がさらに低下して地殻ができ、地殻上で、多くの火山が盛んに噴火を繰り返した。この噴火にともなって、二酸化炭素とアンモニアが大量に放出された。
水蒸気と多少の窒素も含まれていたが、酸素は存在しなかった。
この原始大気は、二酸化炭素が大半を占め、微量成分として一酸化炭素、窒素、水蒸気などを含む、現在の金星の大気に近いものであったと考えられている。 100気圧程度と高濃度の二酸化炭素の温室効果で、地球が冷えるのを防いでいたと考えられている。 #目 次
海の形成の秘密
古い変成岩に含まれる堆積岩の痕跡などから、43~40億年前頃に、海洋が誕生したと推測されている。地球表面が冷えてくると、大気の主成分の水蒸気が雨となって地上に降り注ぎ、海ができる。
年間降雨量が10mを超える凄まじい豪雨が、1千年近くもつづき、原始の海が生まれたとされている。
海ができると、大気中の水に溶けやすい成分は、海に溶け込む。
初期の海洋は、原始大気に含まれていた、火山ガスに含まれている亜硫酸や塩化水素などが水に溶けて硫酸、塩酸になる。

こうして初期の海は、酸性であった。
しかし陸地の金属イオンが雨とともに海に流れ込み、中和されたと考えられている。
中和されると、二酸化炭素が溶解できるようになり、原始大気の半分とも推定される、大量の二酸化炭素を吸収していった。そして、海水中のカルシウムやマグネシウムと反応し、石灰岩(炭酸塩鉱物)となり地殻に固定される。
こうして、大気からは二酸化炭素が取り去られていく。
水蒸気が、紫外線を受けて光解離することで、酸素が生成されてはいたが、鉄などの酸化により、すぐに吸収されたため、大気中に酸素はほとんど残らなかった。
こうした結果、地球の大気は、水に溶けにくい窒素が主成分となった。これは、表面に液体の海を持たない、他の地球型惑星の大気とは対照的である。

惑星の表面に、水が存在できる条件として、恒星に近すぎると熱すぎて蒸発する。恒星に遠すぎると寒すぎて氷となる。
地球は、太陽からの距離が「水が凍らず、沸騰せず、液体を保てる温度帯」であること、そして適度な大気圧があることが、表面に海を保持することが出来るのである。
さらに大気圧が1気圧(1013ヘクトパスカル)のとき、水は0℃〜100℃の温度帯で液体である。大気がもっと高圧であれば、その温度帯は広くなる。大気がもっと低圧であれば、その温度帯は狭くなる。
水は、気圧が高いほど沸点は上がり、氷点が下がる。結果として、液体で存在する温度帯が広くなる。
逆に、気圧が低いほど、沸点は下がって、氷点が上がり、結果として液体で存在する温度帯が狭くなる。 #目 次
太陽と地球の位置の秘密
地球と太陽の距離は、約1億5千万㎞で、光の速度が(真空中で)秒速約30万㎞だから、太陽光が地球に届くのに500秒かかる距離である。仮に、地球と同じ反射率が0・3の物体をおくと、それはマイナス18℃で熱平衡する。熱平衡とは、温度の高い系から、温度の低い系に、熱エネルギーの移動がおこり、何の変化も生じないエントロピー最大の状態をいう。しかし、現実の平均表面温度は、15℃である。
つまり、現実は33℃も温度が高いが、それは、そもそも地球の大気が、「温室効果」を発揮しているからである。

水が存在できる条件として、太陽系では、金星の軌道付近が近い方の限界、火星付近が遠い限界と考えられる。事実、金星にもかつては水があったが、温室効果によって熱暴走を起こして現在の姿になった。大きさや密度が地球とよく似ている金星には、非常に厚い大気があり、そのほとんどが二酸化炭素で、そのため、二酸化炭素の強い温室効果で、金星の表面の温度は、昼も夜も摂氏470度と、太陽により近い水星よりも高くなっている。
大規模な火山活動が約5億年前に起きたと推測されているが、それ以前のクレーターなどに、氷や水が存在した可能性がある。
火星にも、かつて表面に水があった可能性は高いが、現在は、無い。2005年、BBCは火星の北極地方のクレーターで、氷が発見されたと報じた。この円板状の氷は、暗く低温の砂丘の頂上に、薄い層状の霜が凝結してクレーターの底に広がったもの。
火星の赤道付近は日中20℃を越すこともあり、高緯度でなければ氷は存在できない。
また、液体の水も、火星の大気は希薄で、大気中の水蒸気圧が小さいため、火星表面のほとんどの地域で、すぐ蒸発するので存在できない。
地球は、その3分の2を海に覆われた水の惑星である。
水は太陽のエネルギーにより海や地表から蒸発し、雲を形成し、雨や雪となって再び地表や海に降り注ぐ。 地球にはおよそ14億㎞立方の水がある。
そのうちの約97%は海水で、残る約3%の淡水も、その7割は北極・南極地域の氷として存在しているため、地下水を含め川の水や湖沼など、生活に利用できる水は、ほんの0・8%(約1100万km3)にすぎない。 #目 次
大気に酸素生成
32億年前、すなわち地球誕生から凡そ14億年の後、それまでの古細菌と異なる、シアノバクテリア(藍色細菌)と呼ばれる生物が誕生した。細菌の1群であり、光合成によって酸素を生み出す、酸素発生型光合成細菌である。シアノバクテリアは藻類の仲間といわれるが、真核生物の他の藻類とはちがって、細胞内に核がない原核生物(バクテリア)である。
海水に広く分布し、珊瑚礁のようなコロニーをつくり、大繁殖して、地球の光合成生産に大きな貢献をし、現在の大気を作り上げたと考えられている。
シアノバクテリアは、それまでは、嫌気性生物を傷つける存在だった光を、エネルギーとして使い、周りに無尽蔵にある二酸化炭素と反応させることで、硫化水素を使うよりも効率的にエネルギーを得られるようになった。しだいに数を増やし、光合成で大量の酸素を放出するようになり、地球の環境と周囲の生命に多大な影響を及ぼしていく。
自からエネルギーを作り出す仕組みを獲得したことで、生命はより大きなエネルギーを獲得し、細胞を大型化させることが出来るようになる。
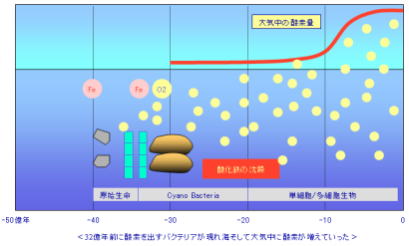
シアノバクテリアは、光合成により効率的に、大きなエネルギーを得る方法を獲得したが、一方で生命にとってはリスクのある方法でもあった。二酸化炭素を光と反応させることで、その廃棄物として酸素が作られる。酸素は、細胞膜や遺伝子を傷つけ、生命に害を及ぼす有毒物質でもある。
それまでに現れた生物は、酸素のない環境でしか生きられない、嫌気性の生物がほとんどあった。
シアノバクテリアによって作り出された酸素は、最初は海洋中に溶け込んだ鉄を酸化させることにより消費された。
27~20億年前の短期間に、海洋中の鉄は酸化鉄となって沈んだ。
現在、鉄鉱層から得られる鉄は、ほとんどがこの時期に作られたものである。
鉄を全て消費すると、次第に海洋中の酸素濃度が上昇した。
酸素と触れた生命は傷つき死滅する。
生物自らが作り出したものにより、それまで過ごしていた環境が激変したのである。環境が変化したとき、その変化に耐えることができない生物は死滅し、絶滅する。
それまでの生物は、酸素という毒に耐性を持ったものはほとんどいなかった。酸素は、この頃の嫌気性の生物にとっては、酸素濃度が上昇することは、とんでもない環境破壊でしかなかったのである。かくして、生命史上最大の危機が訪れる。 #目 次
オゾン層の形成
オゾンは、酸素原子3個からなる気体で、地上から約10~50㎞の成層圏、特に約20~25㎞の高さに多く存在する。このように成層圏内にあるオゾンの層を、一般に「オゾン層」という。このオゾン層は、地球誕生と同時に形成されたわけではない。
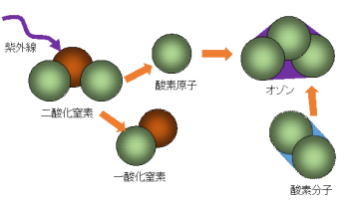
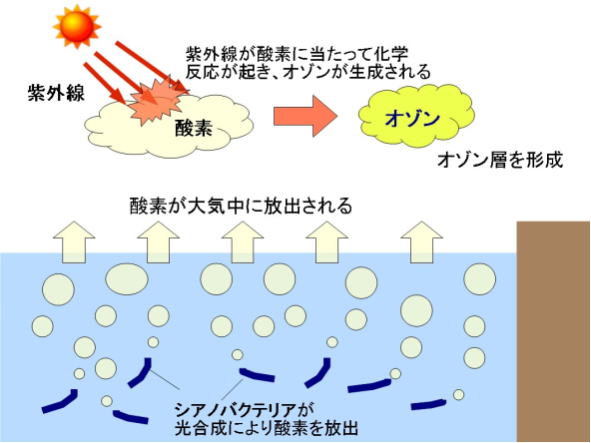
誕生当初の地球の原始大気は、主に二酸化炭素からなり、酸素分子はほとんど存在しなかったため、オゾンもほとんど存在しなかった。約27億年前、光合成により、酸素を放出するバクテリアが誕生して以来、大気中の酸素は増加し、その一部が、太陽からの紫外線で化学反応を起こすことで、オゾンが生成されていった。大気中に酸素分子が増え始めると同時に、オゾンも増え始め、オゾンの量が増えるにつれ、次第に大気上空でオゾン層が形成されるようになった。
原始大気には、紫外線を吸収する物質が無いため、地上まで強い紫外線が降り注いでいた。
その後、酸素濃度が上昇するに従いとオゾンが増え、地上に降り注ぐ紫外線の量は急速に減少していった。しかし当時、オゾン濃度が高いオゾン層が存在したのは、成層圏ではなく、地上付近であった。これは、酸素濃度が薄いため、酸素を光解離させる紫外線が地上近くまで届くからである。酸素濃度が上がると同時に、紫外線の到達できる限界高度が高くなり、これに伴いオゾン層も上空へと移っていった。
原始大気では、酸素濃度の上昇ペースに比べ、オゾン濃度の上昇ペースの方が非常に大きかった。
例えば、酸素が現在の100分の1と薄かった20億年前の大気でも、オゾンは現在の5分の1であった。
オゾンの濃度は、酸素に比べれば非常に薄く、酸素が少ない原始大気でも、紫外線の量は、過去にも大きな変化は無いためで、現在と比べて、それほど少なくない量のオゾンが生成されていた。
また、5億4千万〜5億3千万年前のカンブリア爆発や、4億年前の脊椎動物の陸上進出(両生類の誕生)に関しても、生物に有害な紫外線を低減する、オゾン層との関係が考えられている。
このころは、酸素濃度の上昇によって、オゾン層の高度が高くなり、地上付近のオゾン濃度が低下した時期、およびオゾン濃度が高くなり、地上の紫外線が更に減少した時期に一致する。
ただし、カンブリア爆発の原因を、多細胞生物の接着分子の生合成に必要とされる、酸素濃度の上昇や、浅海域の拡大による、生物の生息範囲の増加に求める説もある。
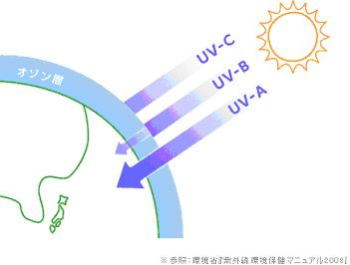
オゾン層は、生物にとって有害な、太陽からの紫外線(UV-B)の多くを吸収する。そのため、海で誕生した生物が、陸上に進出できるようになった。成層圏にあるオゾンの量は、地域や季節によって異なり、一般に緯度が高いほど多く、反対に赤道付近は少なくなり、また春先に増え、秋に減っていく傾向がある。#目 次
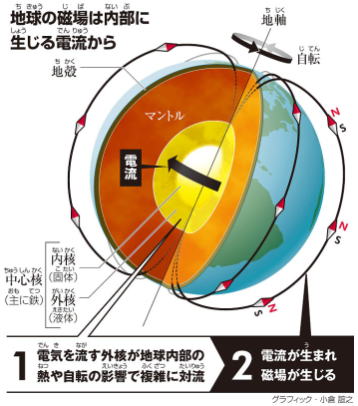
地球磁場の成立
約27億年前ころ、地球環境に大きな変化があった。
地球内部のマントルと核の動きが安定し、中心核の鉄が、磁石のように働き始め、磁気を放出し始め、地球を磁気のバリアが包むようになった。
地球の磁場は、主に地球(電離層等を含む)に流れる電流に起因しているが、地磁気の発生原因は、今でも完全には解明されていない。
ただ地磁気の成因の99%は地球内部にあり、1%は地球外(太陽表面から荷電粒子等)にある。
ガウスは、地磁気のデータから、地球の磁場の成因の99%は地球内部にあることを証明し、80%は双極子(棒磁石)で説明できることを明らかにした。
地磁気は、常に一定ではなく、絶え間なく変化している。その変化の要因は、地球外部要因として、まず太陽フレアの存在である。
磁気嵐や、激しいオーロラが発生したときには、数秒から数日のスケールで激しく変化する。
このような現象は、太陽風と関係がある。
次に太陽放射がある。磁気嵐やオーロラがない場合でも、一日周期で数十 nT(ナノテスラ ―地磁気の大きさの単位、SI単位系の磁束密度の単位) 程度の変化が見られる。
このような1日周期の変化を、日変化と呼ぶ。日変化は太陽放射と関係がある。
地球内部から発生する磁場の変化としては、マグマの活動による地磁気の変化、つまり多くの強磁性鉱物は、キュリー点を超えると、磁化を失う。
つぎに、永年変化がある。磁場の逆転よりも、もう少し変動の振幅が小さい、数年から数千年程度の時間スケールの、磁場変動のことを永年変化と呼ぶ。
地磁気は年々弱くなっており、ここ100年では約 6% 弱くなった。これはあと1千年足らずで、地磁気が消滅してしまうほどの減少率である。ただ、この程度の磁場変動は、過去においてもそれほど珍しいものではない。
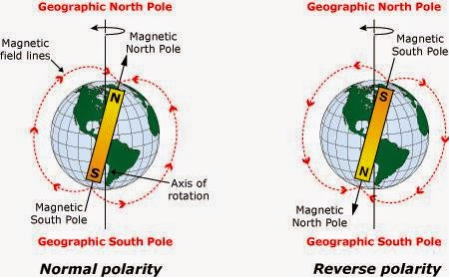
さらに地磁気逆転現象がある。地磁気の逆転は、地磁気極のN極とSが反転する現象である。
古い火山岩などがもつ磁化を測定することで、過去の地磁気の様子を推定するという古地磁気学によって明らかにされた。地磁気は、平均すると 100万年に
1・5 回の割合で逆転を繰り返しているが、その割合はかなり不規則である。
たとえば白亜紀には、1千万年以上にわたり逆転のない期間があったと推定されている。
地磁気は、78万年前にN極とS極は逆転しており、過去360万年で11回逆転していたことが明らかになった。
地球の磁場は、概ね磁気双極子である。つまり、地球の中心に置かれた一つの棒磁石に似ている。
現在は北極部がS極、南極部にN極に相当し、それぞれ北磁極と南磁極と呼ぶ。ただし、非双極子部分は、地球上に「瞳のような形」で存在する。
日本語では「磁極」という語は、一般に磁石の極という意味と、地理極(真の北極や南極)に対する語として、地磁気的な極である北磁極と南磁極の総称としての「磁極」がある。
地磁気の磁力線は、赤道付近を除けば、地面に対して平行ではなく、地面と斜めに交わるかたちになっている。ある地点で、水平面と地磁気のベクトルとがなす角を伏角という。
地磁気が、地面に向かって突き刺さる方向の場合がプラス、地面から出て行く向きの場合が、マイナスとなるように定義される。
伏角は、南半球のほとんどでマイナスで、南の磁極に近づくにしたがって マイナス90度に近づく。
また、北半球のほとんどでプラスとなり、北の磁極に近づくにしたがって プラス90 度に近づく。
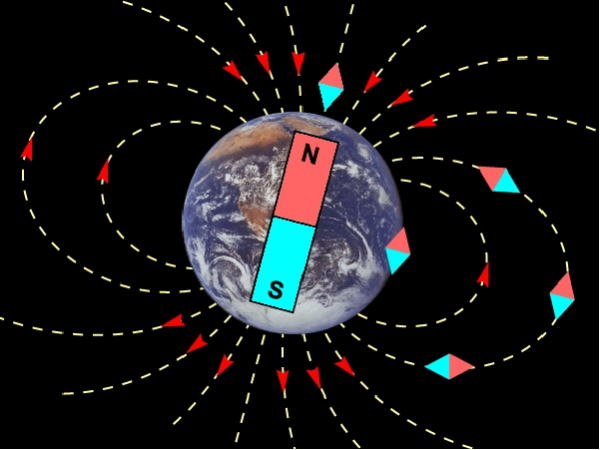
地磁気のベクトルを、水平面に投影したとき、地理上の真北となす角を偏角と呼ぶ。偏角のもっとも大きい要因は、地球の双極子磁場が自転軸に対して傾いていることである。
地球の双極子磁場は、自転軸に対して約 10・2度傾いているため、地理上の極と磁極の位置にはずれがある。
地磁気の極には「磁極」と、「地磁気極(または磁軸極)」という2つの極がある。
北磁極は、方位磁針のN極が真下を向くところで、南磁極は方位磁針のS極が真下を向くところである。現在、磁極は地球の中心に対して対称な位置にはない。
地磁気北極(北磁軸極)、地磁気南極(南磁軸極)は、地球の磁場を磁気双極子としたとき、地磁気の分布が観測された分布図と同じになる、棒磁石の長さ方向への延長線が地表面へ出てくる2地点である。地磁気極は、地球の中心に対して対称な位置にある。 #目 次
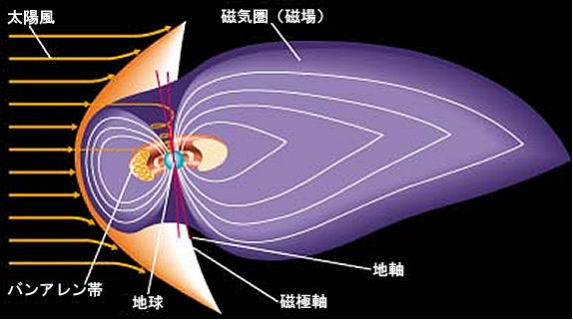
太陽風と磁気バリア
太陽の表面では巨大な爆発を活発に繰り返し、コロナを吹き出している。
その中でも特に巨大な爆発は、太陽の引力を振り切り、超高速で大量の粒子を吹き出す。これが太陽風と呼ばれている。 太陽からはこの太陽風によって、電気を帯びた陽子や電子などの荷電粒子がたえず流れ出している。
この荷電粒子を「プラズマ」、その流れを「プラズマ流」と呼び、オーロラを発生させる元とになっている。
太陽風は約百万度の高温で、電子と陽子が分離してイオン化したプラズマ粒子のガスである。
地球付近に到達した状態では、太陽風は温度が約10万度、速度が秒速450㎞で、プラズマ粒子数が1㎝立方あたり5~10個となっている。
太陽には、地球と同様に固有の磁場があり、吹き出す太陽風は、この磁場を引っぱり出しながら、地球にまで届く。

一方、地球がもつ独自の磁場圏(磁気バリア)は、太陽の磁力線を跳ね返しているので、太陽の磁力線の中を進む、太陽風プラズマは、直接には地球に侵入してこれない。
磁石としての地球磁場は、北極から南極へと磁力線が走る対称な形ではなく、太陽風によって大きな変形を受けている。
太陽風が磁気圏とぶつかる面は、太陽風の圧力と磁気圏の磁場の圧力とが釣り合うまで圧縮されるが、平均的な位置は地球の半径の10倍、約6万㎞のところにある。
太陽風に吹きつけられた磁気圏は、吹き流され、太陽と反対側の地球の夜の側に流される。磁気圏の流されている部分を、「磁気圏尾部」と呼ぶ。この尾の長さは、地球の半径の数百倍、100万㎞以上にも引きのばされていることが確かめられた。
太陽風が繰り出す強烈なプラズマ流から、その地球の生命体を守っているのが、地球の磁場である。しかし、磁場の発生メカニズムについては、よく分かっていないことが多い。
2013年11月、欧州宇宙機関 (E S A) は、 3基の地磁気観測衛星を打ち上げた。更に、デンマーク国立宇宙研究所のグループが、衛星から発信された計測データを解析した結果、驚くべき地球の秘密が明らかになった。
地下3千㎞の外核と呼ばれる層では、液状となった鉄が、大気上空のジェット気流のように対流し、これによって強力な磁場が作られている可能性が高いという。
その流れの速度は、およそ時速約6mである。ジェット気流 (時速 300km以上) に比べると、決して速いスピードではないが、流れは極端に密度の高い液状の鉄である。したがって、その運動量は巨大といえる。
対流を生み出している力は何か。
おそらく、磁場の変化で発生した浮力と考えられている。
このように約27億年前ころ、地球の中心核の鉄が、磁石のように働き始め、磁気を放出し始め、地球を磁気のバリアが包むようになった。

それまで太陽風により、地球まで到達していた生命に有害な荷電粒子(主に陽子・電子)は、磁気圏のバリアに遮られるようになる。このことは、海面近くの光が届く環境でも、生命が存在できるようになることを意味する。それまでは、有害な荷電粒子が届く環境では、生物の遺伝子が傷つけられ、生存し、増殖することができなかった。
海面近くの環境の危険性が低下したことで、光合成を行う生物は、より安全に、より活発に、海面近くで増殖できるようになった。磁気バリアの誕生により、シアノバクテリアの増殖と、地球の酸素濃度の上昇のスピードに拍車がかかった。
磁気圏のバリアで進路を曲げられた電子は、地球の極地から磁場内に進入する。この時進入した電子が、大気中の酸素・窒素原子と反応を起こし、光を放出しているのがオーロラの正体である。
#目 次
最初の陸地
冥王代は地質学的証拠のない時代ながら、証拠の岩石や鉱物という矛盾したものが発見されている。現在までに、多く地域か38億年前より古い岩石や地層が発見されてきた。
地球最古の岩石は、1989年カナダの北西地域のアカスタ地域から、39・8億年前の岩石 ―片麻岩が発見された。片麻岩とは、変成岩の一種で、葉片状または縞状を呈し、花崗岩によく似ていて、主として長石・石英・雲母・角閃石などから成るもの。
それまで、最古の岩石や地層は、グリーンランドの 38億年前のものであった。一気に2 億年も、最古の岩石の記録を塗り替えた。

地球最古の鉱物は、西オーストラリアのジャックヒルズの、クォーツァイトに含まれる、44億年前のジルコン、地球最古の地殻の痕跡はカナダのハドソン地域の片麻岩で、マントルからの分離は42億年前である。
その後、アカスタでは、さらに古い40 億年前の岩石も発見された。アメリカ合衆国、中国、南極など各地から、38 億年前ころの古い岩石が発見されてきた。
冥王代の間に、地球の主な構成要素は、すべて形成されて所定の位置に収まったが、現在とは様相が大きく異なっていた。冥王代、始生代、原生代をまとめて、先カンブリア時代と呼ぶ。非常に稀ながら、45億年前までの岩石は月で発見されている。
#目 次
始生代の地質
始生代とは、地質時代の分類のひとつで、40億年前(または38年前)か25億年前までの間をいう。この時代から、地殻を構成する岩石が見つかりはじめている。
最も古いのは、カナダのスレーブクラトンのアカスタ片麻岩で、約40億年前に形成されたもので、この岩は形成後に激しい変成作用を受けている。このため、当時の地球表層の環境を解読するのは困難である。

アカスタ片麻岩石
当時の地表の状況が判明できる最古の地層は、グリーンランド西部イスア地域の、イスア緑色岩帯で、約38億年前のもの。グリーンランド、カナダ楯状地、バルト楯状地、スコットランド、インド、ブラジル、オーストラリア、南部アフリカなどに残っている岩石のほとんどは、変成作用を受けている。
太古代の岩石は、現在の大陸地殻表面の約4・5%を占めているが、地表に出ていない分まで含めると、現在の約10%とされている。
この時代の陸地面積が、現在より大幅に少なかった可能性が高いが、現在の大陸地殻を構成する岩石(花崗岩類)の、大部分は当時すでに地表に存在し、その後、再溶解してリサイクルされたものであるという説もある。
始生代の終わりの年代は、顕生代のような明瞭な地質学的事項がないため、1981年に提唱された「25億年」が使われている。
地球は45~46億年前に誕生したとされるが、当時は、微惑星の衝突で解放されたエネルギーで、地球内部は現在よりも高温となっていた。その後、地球は徐々に冷却されていく。

38億年前のイスア緑色岩帯
最初の岩石は約40億年前ながら、まとまった地層が世界各地で見つかるのは、38億年前からである。
38億年より前の地層が残っていないのは、現在よりも高温で活発なマントル対流のため、当時形成された地殻はすべて、マントル内部に取り込まれてしまったことが原因とされている。一方、39億年前頃に、地球と月が同時に大規模な隕石衝突を受け、当時の地殻が破壊されてしまったという説もある。
なお堆積岩の分析結果から、30億年より前の海水温度は、60~120℃という高温であったと推定されている。

ストロマトライト
始生代を通じて、大気中には酸素はなく、窒素と二酸化炭素が主体であった。
27億年前頃から、大陸周辺にシアノバクテリアが形成した大規模なストロマトライトから、酸素が放出され始める。その酸素は、縞状鉄鉱床の形成に消費されてしまい、大気中には移行していなかった。ストロマトライトとは、藍藻類とよばれる特殊なバクテリアの群集が、堆積物粒子を結合させたり、炭酸カルシウムを沈殿させたりして作った堆積構造をもつ岩石の総称。
上記の38億年前のイスア地域の地層から、縞状鉄鉱床・炭酸塩岩・枕状溶岩・礫岩層が見られるが、前3つは当時海が存在したこと、礫岩層は陸地があったことを示している。またイスア地域の地質構造は、付加体としての特徴を示しており、当時既にプレートテクトニクスが機能していたと推定される。
35億年前の地層は、アフリカ南部やオーストラリアのピルバラで見つかっている。ピルバラ地域のノースポールからは、35億年前の枕状溶岩の上に載ったチャートの層から、最古の生物痕跡と思われる化石が見つかっている。 #目 次
大陸生成
大陸の地殻を構成する花崗岩の組成は、その下のマントルの組成と大幅に異なっている。海洋地殻を形成する玄武岩は、マントルの一部が溶解してできたもので、花崗岩は玄武岩が水の存在下で、再度部分溶解して生まれる。そのため、地球誕生当初の地表には大陸地殻は無く、その後年代が下がるにしたがって、大陸が増えてきたとされている。

陸地の生成は、一定のペースで進んだのではなく、段階的に起こったとというデータがある。つまり世界各地の花崗岩の中の、ジルコン結晶の生成年代を分析した結果、27億年前と19億年前に、ジルコン生成のピークが認められ、この時期に集中的に陸地が生まれたとされている。
27億年前には、大陸の周辺の浅い海に、大規模なストロマトライト(藍藻 ―シアノバクテリア類の死骸と、泥粒などによって作られる層状の構造をもつ岩石)が形成されたと考えられている。
なお始生代は、マントルの温度が現在よりも高かったため、マントルが部分溶解してできるマグマの成分も現在と異なっていて、マグネシウム分が非常に多いコマチアイトなど、現在のマグマでは見られない成分の、火成岩が存在した。

35~38億年前の地表の状況
また花崗岩も、後世にみられない組成をも ち、ナトリウム成分に富んだトーナル岩・トロニエム岩・花崗閃緑岩からなり、頭文字からTTGと呼ばれている。
マントルの温度が高かったため、沈み込みプレート自体が、比較的浅い地下で融解して大陸地殻に貫入したためと考えられている。 #目 次
原生代初期の地表
27億年前、非常に活発な火山活動で陸地が大幅に増えた。
増えた大陸周辺の浅い海に、光合成を行うシアノバクテリア集合体の、ストロマトライトが大規模に形成された。
ストロマトライトから放出された酸素は海中に拡散し、当時の海中に大量に溶解していた、2価の鉄イオンを酸化し沈殿させ、縞状鉄鉱床を生成した。
縞状鉄鉱床の生成のピークは、27億~19億年前までであった。
始生代の大気には、酸素はほとんどなく、大量の二酸化炭素と窒素が大気の成分であった。原生代に入って、ストロマトライトの活動で酸素が生成され始めたが、浅海に2価の鉄が十分ある間は、酸素が直ちに消費されるため、大気中の酸素濃度は非常に低いレベルのままであった。

その後22億年前頃から、大気に酸素が含まれていたことを示す「赤色土壌」や、「赤色砂岩」が出現するようになった。
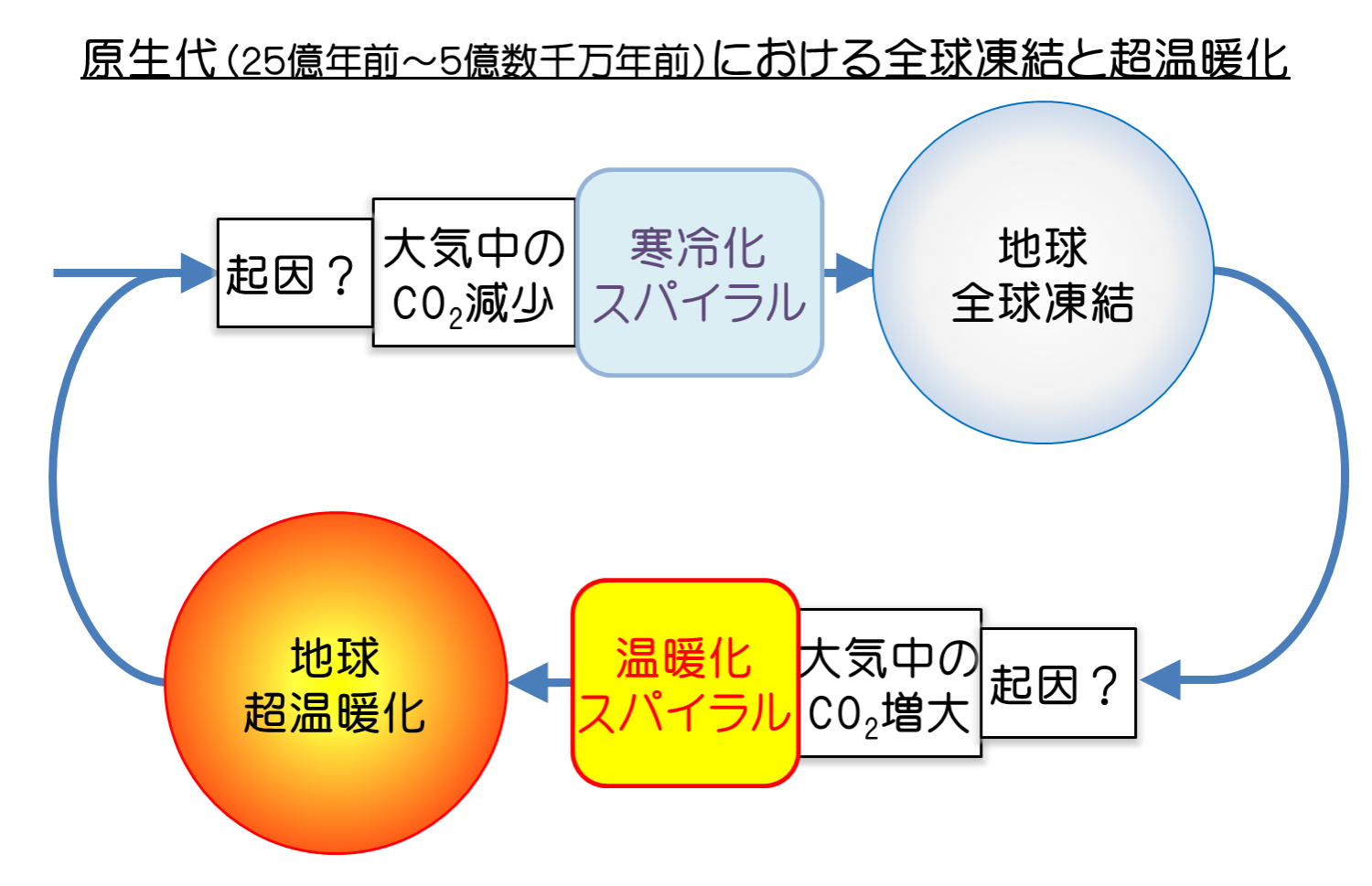
その後、大気中の酸素の比率は徐々に増えてゆく。22~23億年前に、地球は寒冷化し、何回かの氷河時代を迎えたが、最も寒冷化したヒューロニアン氷期には、赤道近くまで氷結し、スノーボールアースとなった可能性がある。
寒冷化の原因は、大気中の二酸化炭素濃度が下がって、温室効果が減ったためと推定されている。二酸化炭素濃度減少の原因は、大陸の拡大によって、岩石の風化量が増え、風化岩石中の金属元素が、空中の二酸化炭素を消費したと考えられる。また、さらに風化した塩類が海に入って、大量の栄養塩類となり、生物活動(光合成)を活発化させ、二酸化炭素を消費したことも考えられる。 #目 次
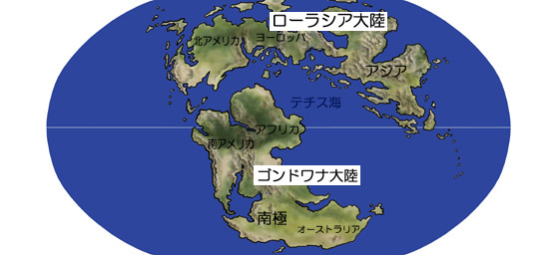
超大陸の形成― ロディニア大陸
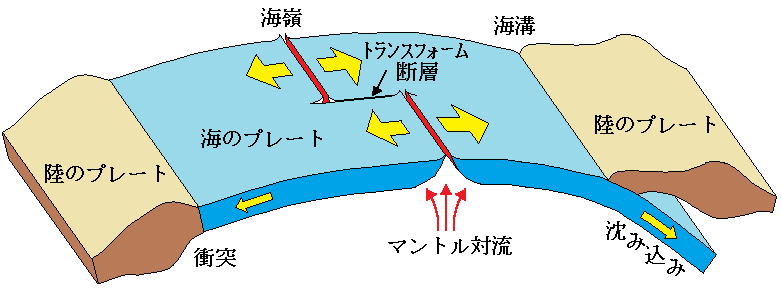
プレートテクトニクス(大陸や海底プレート相互の変動を、プレートの水平運動によって理解する学問)では、プレートが動くことで、その上の陸地も地表を移動する。
地球の歴史では、殆どの大陸が1か所に集結して、巨大な超大陸を形成したことがあった。顕生代に存在した「パンゲア」は有名だが、原生代後期の10億年前頃にも、「地上のほとんどの陸地が集まった「超大陸」が存在した、とする研究結果が報告されている。この超大陸は、「ロディニア大陸」と呼ばれ、約7億年前に3つに分裂したとされている。
ロディニアは、それ以前にあった比較的大きなヌーナ大陸、コロンビア大陸、アトランティカ大陸の、3つが合体したものである。
これら3大陸は、約19億年前にあった活発な大陸成長のピーク期に、もっと小さな陸地が集合し成長して生成したとされている。
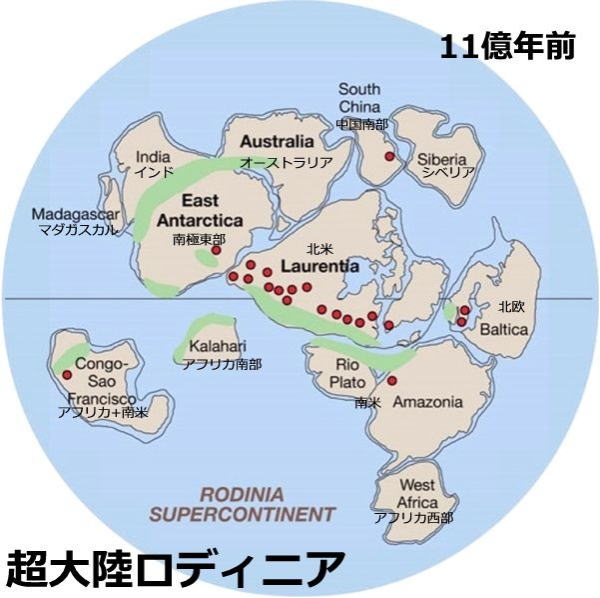
20世紀後半の研究の進歩で、過去の大陸移動の様子が詳しくわかってくると、パンゲア大陸以前にも、超大陸が存在したことが分かってきた。ロディニア大陸は、グレンヴィル造山運動で形成されたと考えられているが、その時期については、研究者間でもまだ見解が一致していない。
1991年、ポール・ホフマンが、科学雑誌『サイエンス』に、ロディニア大陸の主要な配置について発表した。 ロディニア大陸は、パンゲア大陸が形成された地域からほぼ正反対の、現在の太平洋地域に、やや南半球寄りに形成されたと考えられている。
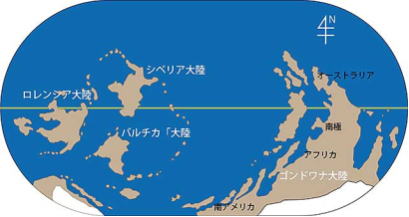
ロディニア大陸が分裂 右ゴンドワナ大陸
ロディニア大陸があった時代に、全球凍結が起きたという説がある。その原因のひとつは、当時ほとんどすべての陸地が低緯度にあり、南北両極ともに海だったことだという。陸地は海よりも熱の反射率が高く、赤道近くに陸地が多いほど、太陽エネルギー吸収の効率を下げることになる。
ロディニア大陸が分裂した後は、ゴンドワナ大陸と呼ばれるかなり大きな大陸と、シベリア大陸、ローレンシア大陸、バルティカ大陸と呼ばれる、小さな大陸へと分裂したと考えられている。21世紀初頭では、ロディニア大陸よりもさらに前の超大陸として、約10億年前に存在した、パノティア大陸の存在が指摘されている。あるいは約10億年前に存在した超大陸をロディニア大陸、約7億年前に存在した超大陸をパノティア大陸と呼ぶこともある。#目 次
プレートテクトニクス
顕生代の地球環境は、地球外からの影響の第一として、太陽からの光エネルギー到達量がある。太陽は誕生以来、徐々に明るくなってきており、顕生代では地球が受け取るエネルギー量は、1億年で約1%ずつ増加している。
その他の地球外要因として、他の天体との衝突がある。
地球には常時、小さい隕石が落下しており、ごくまれに大きな隕石も落下している。地球に衝突する天体の大きさと頻度は、反比例関係にあり、バリンジャー・クレーターを生成した大きさの隕石は、数千年に1回程度、恐竜絶滅の原因の可能性が検討されている。K―T境界では、直径10~15㎞レベルの衝突は、約1億年に1回程度衝突すると考えられている。
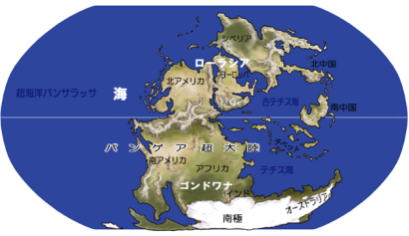
顕生代が始まる直前の5億5千万年前、いくつかの大陸が海洋を隔てて存在する超大陸パンゲアが分裂して、大西洋やインド洋が生まれ、現在の大陸分布になった。
大陸と海洋の関係については、プレートテクトニクス理論に基づいた研究によって、大陸の離合集散が明らかになってきた。
また非常に規模の大きな洪水玄武岩と呼ばれる噴火が、顕生代にしばしば発生していることがわかってきた。
大陸と海洋の配置は、顕生代を通じて大きく変化した。
原生代後期に、超大陸ロディニアが形成されたが、この超大陸はすぐに分裂した。
顕生代初期に、分かれていた大陸が集まり始めた。ゴンドワナ大陸と、北米大陸やバルチカ大陸(現在のヨーロッパの一部)は、広い海で隔てられていた。
シルル紀からデボン紀にかけて、古生代の造山運動があり、陸地が増加し、ゴンドワナ大陸は、赤道から南極まで広がっていた。
古生代の後期には、当時存在した全ての大陸が陸続きとなって、超大陸パンゲアが形成された。
古生代最後のペルム紀末の(P-T境界)で、陸上では顕生代史上最大級の、400万立方㎞以上の溶岩流出事件、シベリア洪水玄武岩が発生した。
中生代に入ると、パンゲア大陸は分裂を開始した。 三畳紀末には、アフリカ大陸と、南アメリカ大陸が分かれ始め、その際に割れ目に沿て洪水玄武岩の噴出があった。
この噴火による玄武岩台地は、割れ目となった大西洋をはさんで、南アメリカ大陸とアフリカ大陸の両方に残っており、中央大西洋マグマ区と呼ばれている。
ゴンドワナ大陸は分裂を続け、インド・オーストラリア・南極の各大陸が分離し始める。白亜紀の約1億2千万年前に、シベリア洪水玄武岩をしのぐ規模の、洪水玄武岩の噴火が太平洋の深海底で発生した。
現在、オントンジャワ海台と呼ばれている、玄武岩地形は面積2千万平方㎞、噴出した玄武岩量は6千万立方㎞、または8千万立方㎞とされているが、殆ど全てが太平洋の水面下に存在している。
ゴンドワナ大陸から分かれた南極大陸は、南下して南極に位置した。
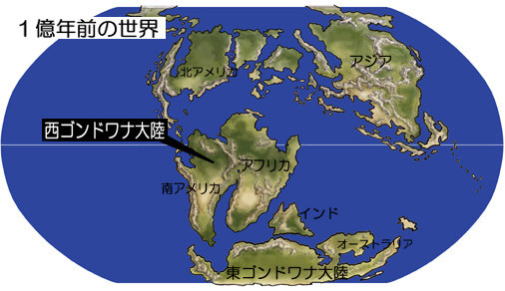
インド大陸は、北北東へ移動してゆき、アフリカ大陸との間にインド洋が開いてゆく。 白亜紀は、温暖な気候と活発なマントル活動の影響で、海面水位が現在より約200m上昇して、陸地面積は減少した。
約6千600万年前の白亜紀最後に、インド大陸でデカン高原を形成する洪水玄武岩の噴火があり、推定約100万立方㎞から250万立方㎞の玄武岩溶岩が噴出した。
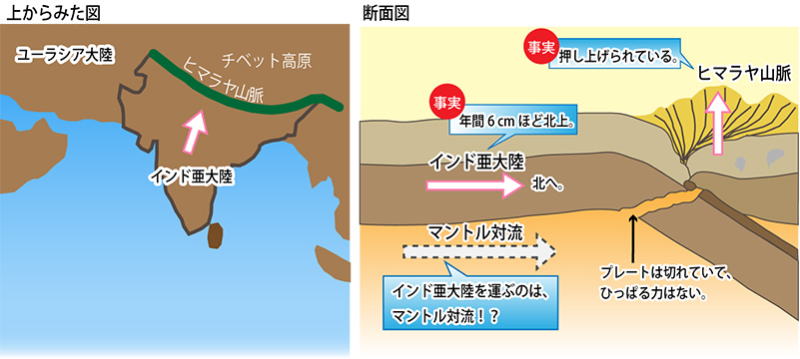
新生代に入ると、北上を続けていたインド大陸が、約4千万年前にアジア大陸に衝突、ヒマラヤ山脈やチベット高原の上昇が始まる。
約3800万年前に、オーストラリア大陸と南極大陸が完全に分離し、約2千万年前には、南アメリカ大陸と南極大陸も離れ、南極大陸が完全に海で囲まれる。
インド大陸は、アジア大陸に衝突したあとも北上を続け、アジア大陸の内部に約2千㎞も突入したため、衝突地点のヒマラヤ山地や、背後のチベット高原は、その下にもぐりこまれたインド大陸に押し上げられ隆起した。
隆起しつつあるヒマラヤ山脈では、高山に対する激しい浸食による、岩石の風化が継続している。約350万年前に、南北アメリカ大陸の間に、パナマ地峡ができて、大西洋と太平洋が分離された。#目 次
プルームテクトニクス
プルームテクトニクス は、90年代以降の地球物理学の新しい学説である。地球マントル内の、大規模な対流運動をプルーム (plume) と呼び、この変動を研究するため、プルームテクトニクスと命名された。
プレートテクトニクス理論が、地球の表面に存在するプレート(厚さ約100㎞)の変動(テクトニクス)を扱うのに対し、この説では、深さ2千900㎞に達するマントル全体の動きを研究する。
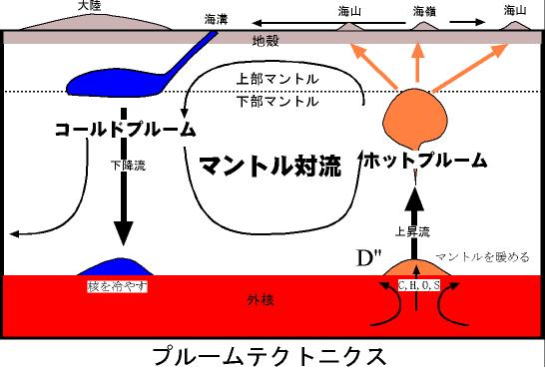
プ ルームとは、羽毛のように舞い上がる「煙」を意味する。
マントルは、半径約6千357㎞の地球の中で、深さ数十㎞~約2千900㎞までの範囲を占めている。その中を下降する、プルームの「コールドプルーム」と、上昇するプルームの「ホットプルーム」が存在する。
プルームの上昇・下降とも、通常時は深さ670㎞の所でいったん停滞する。この部分は、上部マントルと、下部マントルの境目に当たり、マントルを構成する鉱物が、この位置の温度と、圧力を境に相変化するため、この上下でマントルの密度や固さが大きく変化する。 ―と想定されている。
プルームが深さ670㎞付近を超え大きく上昇、あるいは下降したものをスーパープルームという。
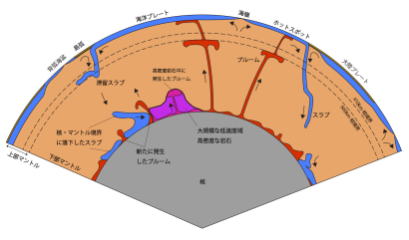
プレートテクトニクスでは、大陸プレートや海洋プレートの動きから、地球表面で発生している造山運動・地震・火山などの説明に至ったが、プレートが移動する方向について検討されておらず、超大陸の形成や分裂を説明することはできなかった。
プルームテクトニクスは、これに説明をもたらした。
また生物の大量絶滅の原因についても、地球内部の動きに起因する大陸の離合集散や、大規模な火山活動による二酸化炭素濃度の上昇に端を発する気候変動と関連付けて、学際的な研究が行われている。#目 次
スーパーホットプルーム
ホットプルームとは、コールドプルームと逆に、深さ2千900㎞の核との境目で、核の熱を受けて高温になったマントル成分が上昇するものをいう。
現在は、アフリカ大陸の下と、南太平洋にスーパーホットプルームが存在し、大地溝帯(グレート・リフト・バレー)が形成された原因で、南太平洋に点在する火山の源であると考えられている。
ホットプルームもまた、外部マントルと内部マントルの境目の深さ670㎞の部分に一旦滞留するため、通常では地上へ激甚な影響を与えることはない。
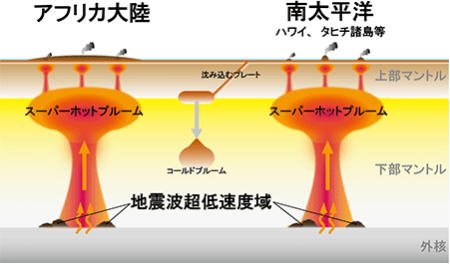
しかし大規模なスーパーホットプルームが、直接地表に達すると、非常に激しい火山活動が発生すると考えられる。
地球生命史上、最も大きな大量絶滅が発生した、2・5億年前のペルム紀/三畳紀境界(P-T境界)では、史上最大級の溶岩噴出事件により、シベリア台地玄武岩(洪水玄武岩)が形成された。
これはスーパーホットプルームによるものと考えられる。
この時期は、超大陸パンゲアが分裂を開始した時期に相当し、プルームの地表への到達と、大陸分裂について相関性が指摘される。 将来的には、次の超大陸ができたときに、直接地表に達する大規模なスーパーホットプルームが、起こると考えられている。現在、次の超大陸形成には、約2億年後に出現すると考えられる ―アメイジア大陸説と、約2億5千万~4億年後にかけて出現すると考えられる ―パンゲア・ウルティマ大陸説がある。
アメイジア大陸説では、現在の太平洋、パンゲア・ウルティマ大陸説では、現在の大西洋での大規模なスーパーホットプルームの発生が推測される。 #目 次
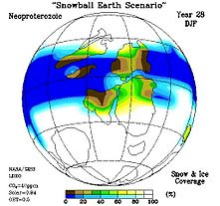
スノーボールアース
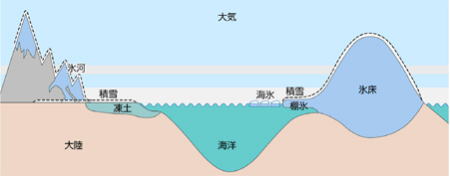
スノーボールアースとは、雪球地球、全球凍結状態の地球のことで、地球全体が赤道付近も含め、完全に氷床や海氷に覆われた時代のことである。
地球はその誕生以来、何度か「氷河期」と呼ばれる寒冷な気候に支配される時代があった。現在判明しているもっとも古い氷河期は、南アフリカで発見された約29億年前のポンゴラ氷河時代で、最も新しいものは、現在も続いている「新生代後期氷河時代」である。
最近の約1万年は、氷河期の中で比較的温暖な間氷期とされる。

ところが「原生代初期」の、ヒューロニアン氷河時代(約24億5千万年前から、約22億年前)の最終期と、「原生代末期」のスターチアン氷河時代、およびマリノニアン氷河時代(約7億3千万年前~約6億3千500万年前)に、地球表面全体が、凍結するほどの激しい氷河時代が存在した。
―という考え方が、地球史の研究者の間で主流となっている。これを「スノーボールアース仮説と」いい、1992年にカリフォルニア工科大学のジョー・カーシュヴィンク教授が、アイデアとして専門誌に発表したのが発端である。
その後98年、ハーバード大学のポール・ホフマン教授が、南アフリカのナミビアでのキャップカーボネイト調査結果などをまとめて、科学雑誌サイエンスに投稿し大きな反響を得た。

この仮説で注目するべき点は、「ありえない」と考えられてきた、「全球凍結」という壮絶な環境変動が、実際に起こったらしいこと。
それが原因となって、原生生物の大量絶滅(大絶滅)と、それに続く跳躍的な生物進化を、もたらしたとされることであろう。
たとえば、酸素呼吸をする生物の誕生や、エディアカラ生物群と呼ばれる、多細胞生物の出現などが、スノーボールアース・イベントと密接に関わっていると考えられている。
これまでは、地球は形成直後のマグマオーシャンに覆われた灼熱の状態から、徐々に冷え、温暖な気候の時期と、氷河期を経ながら現在に至ったもので、この間に、地球全体が赤道に至るまで、完全に凍結したことは、1度もなかった。 ―と考えられてきた。 その理由は、太陽光を熱源とする「熱収支」を考慮した結果である。
仮に地球全体が凍結したならば、地表はすべて白い氷雪で覆われてしまい、太陽光エネルギーの大半を、宇宙空間へ反射してしまう。この場合、地表温度はさらに低下する。 その結果、地球全体が一旦凍結したならば、現在も凍結しているはずであるという仮説である。

それ故に、現在の地球が温暖な気候で、液体の海をたたえていることが、そのまま、地球が凍りついたことが一度もない。 ―ということの証拠であるとされていた。
また地球生命が、約38~40億年前の誕生以来、ずっと継続していることが、全地球完全凍結という事態が起こらなかった証拠と考えられてきた。
仮に全球凍結状のようなことが起これば、生命がそれを生き延びたとは考えにくく、再び温暖になったときに、生命も再び誕生したと考えるのが妥当とされていた。
スノーボールアース仮説では、地球が完全に凍結したとしても、再び温暖な環境を取り戻す過程を提示し、スノーボールアース状態が存在する可能性を示した。
凍結から脱する要素として、火山活動に由来する、二酸化炭素などの温室効果ガスの蓄積を挙げている。
現在のような地球の液体の海は、大気中の二酸化炭素を吸収するため、大気中の温暖化ガスの濃度は、ある程度に抑えられ、温室効果による温度上昇も抑制される。
しかし、全球凍結状態では、海が凍り付いてしまうことから、二酸化炭素をほとんど吸収せず、火山から放出された二酸化炭素は、海に吸収されることなく、大気中に蓄積していく。
このため、二酸化炭素の濃度は約2千年間かけて、最終的に現在の400倍程度に達したとされる。
その大きな温室効果が、大気の温度を最大で100℃ 近く上昇させ、結果として平均気温は40 ℃ 程度となって氷床が溶けだし、全球凍結状態を脱出したと考えられている。
また生物についても、凍結しなかった深海底や、火山周辺の地熱地帯のような、一定の温度が保たれる場所で、生きながらえてきたと考えられている。 #目 次
スノーボールアースの推移
原始地球の大気の主成分は、二酸化炭素と窒素が主体であった。
この大量の二酸化炭素が、地殻が冷めることで地殻に固定され、大気中の二酸化炭素量が低下する。
すると、温室効果の減少で、地球全体の寒冷化が始まり、極地から次第に氷床が発達していった。氷床が拡大することで、その分だけ太陽光を反射するため、太陽熱が地表に届かず、一層の寒冷化を招いた。その結果、一度加速した寒冷化は止まらず、最終的に、厚さ約1000mにも及ぶ氷床が全地球を覆い、スノーボールアースに至った。

この状態は、数億年~数千万年続いたとみられる。凍結しなかった深海底や、火山周辺の地熱地帯では、わずかながら生命活動が維持されていた。
凍結中も続いた火山活動で、二酸化炭素の供給は続けられ、大気中の二酸化炭素濃度が高まっていった。地表が凍結している間は、岩石の風化も凍結状態だった。
大気中の二酸化炭素濃度が、一定比率に達すると気温が上昇し、一気に氷床の解凍が始まった。短くて数百年単位で、極地以外の氷床が消滅し、気温は約40℃まで上昇したと推定される。
温暖化した気候の影響で、大規模な嵐や台風が頻発するようになり、岩石の風化が促進され、大量の金属イオンが海に供給された。 また長年堆積していた海の沈殿物が、嵐により撹拌され、沈殿物が海の表層部に舞い上がった。
大気中の高濃度の二酸化炭素は、海中に溶け込み、一部は金属イオンと結合し、大量の炭酸塩岩を海底に沈殿させた。
海の表層部に舞い上がった大量の沈殿物や、陸地から供給される栄養塩類が、光合成単細胞生物に利用され、光合成を激しく促した。
またスノーボールアース以前の、光合成生物の酸素放出速度より、遥かに速いスピードで酸素が放出され、大量の酸素が地球に蓄積していった。
スノーボールアース中に、極低温により生物の大量絶滅が起こっていた。しかし、スノーボールアースの終了後、生き残った生物の適応拡散が起こった。

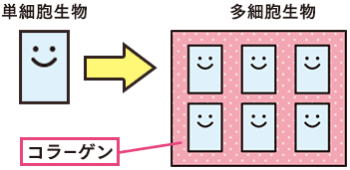
原生代初期のスノーボールアースでは、酸素呼吸をおこなう真核生物の繁栄がはじまった。原生代後期では、一部の生物が、海中の高濃度の酸素を利用し、細胞接着物質であるコラーゲンを産生することに成功する。単細胞間の接合が促進され、多細胞生物が出現するようになった。
原生代後期の、スノーボールアースが始まる前(10億年前)の生物界は、単細胞生物が主体で、多細胞生物は、小形の菌類などがようやく出現し始めた段階であった。
しかしスノーボールアースが終了した、原生代末のエディアカラ紀(6・2~5・5億年前)には、「エディアカラ生物群」と呼ばれる、大形生物が出現している。
大きなものでは長さ1mを超える生物化石が、オーストラリア南部のエディアカラ丘陵から産出した。この突然の大形生物出現と、スノーボールアースの関係について検討が行われている。なお生物の進化は加速し、その次のカンブリア紀には、バージェス頁岩化石に代表される多様な生物群が生まれた(カンブリア爆発)。#目 次
寒冷気候と温暖気候の循環
地質時代区分の「原生代」の後の「顕生代」は、約5億4200万年前から現在までの期間をいう。顕生代とは「肉眼で見える生物が生息している時代」という意味である。
この顕生代の地表の温度を決定付けてきたのは、「太陽から到達するエネルギー量」と、「温室効果ガスの存在量」のバランスであった。
「太陽から到達するエネルギー量」は1億年で約1%ずつ増加するという非常にゆっくりした変化である。
地球の平均気温に対する、温室効果ガスの影響は非常に大きく、現在の地球で「温室効果ゼロ」を仮定した場合の、地表温度はマイナス18℃で、実際の平均気温15℃との間に33℃の開きがある。
この差が、大気中に存在する水蒸気や二酸化炭素などの、温室効果ガスの効果である。
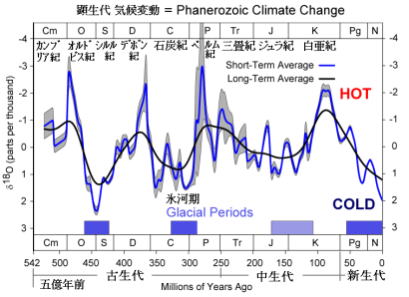
このように顕生代の気候は、大気中の「温室効果ガス」の存在量に大きく左右されている。メタンも温室効果ガスだが、大気中に放出されたメタンは、容易に酸化され、二酸化炭素と水に変わる。このため、長期の温暖化効果としては二酸化炭素の影響が大きい。
顕生代の気候の大まかな特徴は、約3億年周期のサイクルで「寒冷気候」と「温暖気候」が繰り返された。この周期は大陸の集合と分裂の時期と一致している。
つまり大陸が集合する時期に気候が寒冷化し、大陸が分裂する時期に温暖期となった。大陸が分散していた、オルドビス紀からデボン紀までが温暖時期(オルドビス紀末に一時的な氷河期があった)。超大陸パンゲアができ始めた、石炭紀から三畳紀の始めまでが寒冷期で、パンゲアが分裂し始めた、三畳紀末から新生代の初めまでが温暖期にあたる。
アジア大陸にインド大陸が衝突し、アフリカ大陸もヨーロッパに衝突しつつある現在は、寒冷期となっている。
寒冷期と温暖期が発生する原因として、大気中の二酸化炭素の増減による、温室効果の差が挙げられる。
温暖期は火山活動が活発で、マントル内部からの二酸化炭素供給が多く、温室効果が高かった時期に相当する。
大陸が集合・衝突する時には、ヒマラヤ山脈やアルプス山脈のような大山脈ができる。高山は平地より浸食を受けやすく、特にヒマラヤ山脈では多量の雨による激しい侵食を受け、その結果、多量のカルシウムイオンが海洋に供給される。
このカルシウムイオンが、効率的に二酸化炭素を吸収して石灰岩となり、大気中から二酸化炭素を取り除き、二酸化炭素による温室効果を削減するため地球が寒冷化する。顕生代の生物は、この気候変化の影響を受けて進化してきた。
なお現在は、南極やグリーンランド上に、広大な大陸氷河が形成されているため、氷河時代に分類されるが、氷河時代の中では、比較的温暖な間氷期に相当する。
#目 次
氷河期
氷河期は、地球の気候が長期に寒冷化する期間のことで、極地の氷床や、山地の氷河群が拡大する時代で、氷期とも呼ばれる。氷河学的には、氷河期という言葉は、南半球と北半球に、氷床がある時期を意味する。
この定義によれば、グリーンランドと南極に、氷床が存在する現代も、未だ氷河期の中にいることになる。

過去数百万年に関しては、氷河期という言葉は一般的に、北アメリカとヨーロッパ大陸に氷床が拡大した寒冷期について用いられる。アジア地域は氷床が発達せず、寒冷な地帯であったらしい。
この意味で言えば、最後の氷河期は、1万年前に終了したということになる。この約1万年前に終わった出来事を、文献によっては「最後の氷河期」と記載していることもある。科学者の多くは、氷河期が終わったのではなく、氷河期の寒い時期「氷期」が終わったとし、現在を氷期と氷期の間の「間氷期」としている。そのため、最終氷期終了後から現在までの期間を、後氷期と呼ぶこともある。

過去数百万年は、4万年から10万年の周期で、多くの氷期が起こり、これについては研究がさかんに行われている。
各氷期と間氷期では、それぞれ平均気温が異なり、最近の氷期では、年平均気温で7~8℃以上、低下したというデータもあるが、「気温何度から氷期」というわけではない。その間にも小氷期、小間氷期が認められる。
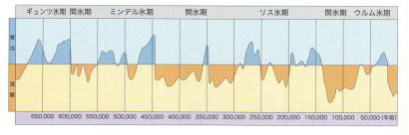
ヨーロッパでは、古い方から「ギュンツ」、「ミンデル」、「リス」、「ウルム」の4氷期に区分されている。数万年単位などの短期的視野ではなく、さらに大局的・長周期的に見ると、過去地球上では、少なくとも4回の大氷河期があった。
この長い「氷河時代」が重要なのは、人類の進化に並行しているからである。
氷期が訪れると、海岸線が極端に遠のき、陸上の大部分が氷に覆われる。そのため動植物も激減し、動植物を食料とする狩猟採集生活の人類にとっては、大きな打撃であった。人類(猿人)になる前は、樹上生活であったが、氷期の環境で、地上生活を始めて2足歩行を開始し、人類となったというのが通説である。
#目 次
過去の大きな氷河期
過去、地球上では、少なくとも4回の大きな氷河期があった。
24億年前から21億年前頃の原生代初期に、最も古い氷河期 ―ヒューロニアン氷期があったことが仮説として考えられている。
証拠が残っているもので一番古いのは ―原生代末期の7億5千万年前からの氷河期で、スターティアン氷期、およびマリノア氷期の6・4億年前である。
これは過去10億年のなかで、おそらくもっとも厳しいものであり、氷が赤道まで覆いつくし、スノーボールアース(全地球凍結、全球凍結)を作り出した。この氷河期の終結が、カンブリア爆発の原因になったと言われている。
古生代には、4億6千万年前から4億3千万年前にかけて、小さな氷河期 ―アンデス−サハラ氷期があり、同じく古生代の3億6千万年前から2億6千万年前にかけても、氷河の拡大期 ―カルー氷期があり、このときには生物の大量絶滅が起きている。
現在の氷河期は、4千万年前の南極の氷床の成長により始まり、300万年前から起きた、北半球での氷床の発達とともに規模が拡大した。更新世に向かうにつれて更に激しくなり、その頃から氷床の拡大と後退の繰り返しによる、4万年と10万年の周期が、世界中で見られるようになった。最後の氷期 ―最終氷期は約1万年前に終った。
#目 次
氷河期の原因
なぜ「氷河期」が起こるのか。
これは大きなスケールで起こる氷河期についても、氷河期の中で起こるより小さな氷期 ―間氷期の繰り返しについても、いまだ議論されている問題である。
一般的な総意としては、大気組成 ―特に二酸化炭素とメタンのフラクションと、「ミランコビッチ・サイクル)」として知られる、太陽を回る地球の軌道要素、そして大陸の配置の組み合わせ、の3つの要素が組み合わされたものがその原因とされている。
・大気組成の変化
3つの要因のうち、「大気組成の変化」は、特に最初の氷河期について重要な原因とされている。スノーボールアース仮説では、原生代後期の、大規模な氷河時代の始まりと終りは、大気中の二酸化炭素濃度の、急激な減少と、急激な上昇が原因であるとされている。
氷河期の大陸の配置
・大陸の配置
北極圏と南極圏に、大陸がどれだけ配置されているかが、氷河期が起こる際に重要であることがわかってきた。
特に、新生代氷河期が始まった原因は、大陸の配置の変化によるところが大きいとされる。それは、大陸の存在によって、寒冷期に雪や氷が集積することが可能になり、この現象は、アルベド効果(陽光を反射し気温をさげる)のような、正のフィードバック効果の引き金となるからである。
また、大陸の配置は海洋や大気の循環システムにも大きな影響を与える。
・地球軌道要素の変化
地球軌道要素は、長期にわたる氷河期では、大きな原因とはならないが、現在の氷河期の中で交互に起こっている、凍結と溶解の繰り返しのパターンを支配していると考えられている。地球軌道と、アルベドの変化の複雑なパターンによって、氷期と間氷期の二つのフェーズが起こるようである。
氷河期については、現在の氷河期、特に最近40万年間について、詳しく研究され理解が進んでいる。
最近40万年のデータは、大気組成や気温、氷床量の指標が記録されている氷床コアの分析から、得られている。
この期間は、氷期 ―間氷期の繰り返しが、ミランコビッチの提唱した周期(ミランコビッチ・サイクル)とよく呼応しているから、その説明として軌道要素が一般的に受け入れられている。
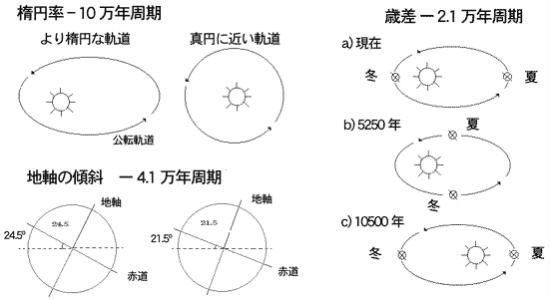
太陽からの距離の変化 ―軌道離心率、地軸の歳差運動(傾いている地球の地軸は、公転面に垂直な方向に対して半径約23・4度の円を描くように移動)、地軸の傾き ―傾斜角が複合して、地球が受ける日射量の変化に影響を与えている。
特に重要なのは、季節性に強い影響を与える地軸の傾きの変化である。たとえば、北緯65度における7月の太陽光の入射量は、計算によれば最大で25%(1平方mたり400Wから500W変化するとされている。
夏が涼しい時、前の冬に積もった雪が溶けにくくなるので、氷床は前進するというのは広く考えられていることである。
日射量のわずかな変化は、「前の冬の雪が完全に溶解する夏」と、「次の冬まで溶けずに残る夏」の間のバランスを調節する。
何人かの研究者は、軌道要素は、氷期の開始の引き金になるには弱過ぎるとしているが、二酸化炭素のような、フィードバック機構でそれは説明できるとしている。
#目 次
海水準の変化
海水準変動という言葉は、主に地質時代の世界的な海水準 ―陸地に対する海面の相対的な高さの変化をいう。
海水準が上昇することは海進、下降することは海退と呼ばれる。
古気候の研究者は、現在を氷河期とみなし、地質時代の中でも海水準は比較的低く、海水準の上昇と低下が頻繁に起こる時代と認識している。
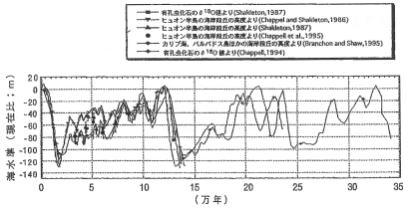
35万年間の海水準グラフ
過去の記録から、およそ1万8千年前の最終氷期最盛期から、6千年前までの間にかけて、海水準が120m以上も上昇したことが分かっている。3千年前以降19世紀までの海水準の変動率は、ほぼ一定で年間で、年間0・1~0・2㎜程度であったが、1900年以降は年に1~3㎜と上昇している
顕生代の海水準の変化は、主に気候が影響している。
すなわち氷河期には、大量の水が氷床として陸上に固定されるため、海水準が低下し、温暖化によって海水準は上昇する。 急激な海水準の低下は、浅海に住む生物の生存に打撃を与え、絶滅の原因となる。
古生代の石炭紀後期から、ペルム紀中期にかけて、地球は寒冷な氷河期(ゴンドワナ氷河時代)であった。パンゲア大陸の南部を形成する、ゴンドワナ大陸が広い範囲で氷床に覆われた。
これらのことから、P―T境界の大絶滅の原因として、海水準の低下を指摘する説もあった。しかし90年代以後の中国南部や、他の地域での研究結果から「P―T境界の50万年前から、海水準は上昇しつつあった」とされている。
地球規模の平均海水準は、固体である地殻表面の形状と、液体である全海水の体積により定まる。そして全海水の体積変化は、全海水量の変化と、温度による体積変化 ―すなわち熱膨張の効果の二つからなる。
全海水量の変化は、主に陸上の氷との相互変換による。
流氷や北極海の氷山のように海面に完全に浮いている氷は、融解しても全海水の体積を変えないので、正確には全海水量とは、液体である海水と、完全に浮いている氷との合計量となる。
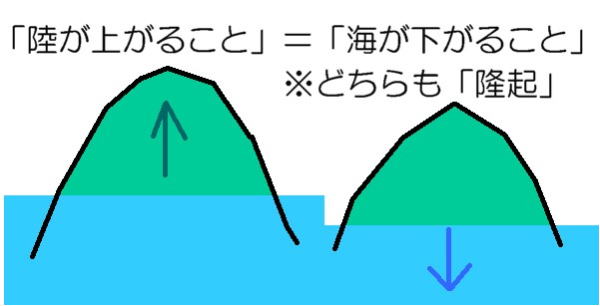
長い時間軸では、海洋の形の変化と陸海分布も、海水準を決める要因になる。また地球内部から地表への水の出入りによる、地表水の全質量の変化が要因となることもある。地球の長い歴史の間では、大陸移動により陸の配置が現在とはかなり違った形状であった。
またプレートテクトニクスの観点からは、海洋プレートの生産が活発になると、海洋底が拡大するため ―現在のインド亜大陸とユーラシア大陸のように、大陸同士の衝突が起きた場合、海洋の面積が増え、相対的に海水準が低くなるという説もある。
大地殻が極付近に大量に集まった時代は、堆積物の記録から、氷期の間著しく海水準が低かったことがわかる。 これは極域の陸上には、雪や氷が集積できるからである。
陸塊が赤道付近に密集している時期には、氷期(寒冷期)が海水準に与える影響は小さい。しかしほとんどの地質時代、長期間の平均的な海水準は現在より高い。
例えば、白亜紀には、現在よりも200m以上高い時期があった。 現在よりも平均的な海水準が低かったのは、2億5千万年前のP-T境界(ペルム紀/三畳紀境界)付近の間のみである。
過去数百万年の新生代氷河期の、氷期と間氷期サイクルでは、海水準は100m以上の変動を示している。
主に海から蒸発した水が、ほとんどは北半球で、氷床として固定され、成長する時に、海水準が低下、後退するときに、融解水が海洋に供給されることで海水準が上昇するからである。現在のグリーンランドと南極の氷床が溶けると、海水準はおよそ80m上昇すると予測されている。
#目 次
生命の定義
そもそも生命と、物質とは何が異なるのか。
生物学での生命の定義は、その内部での物質交換と、外部との物質のやりとり(代謝)、および同じ型の個体の再生産(遺伝と生殖)にある。そのような性質を持つ、最小の単位が「細胞」であり、細胞を生命の最小の単位と見なし、それから構成されるものに生命を認める。 ―というのが一般的である。
生命の特徴・定義を生物学的に整理すると、以下の条件を満たさねばならない。
①自己複製
生命の特徴のひとつに、自己と同じ子孫を複製し増殖する能力持つ事である。
これは核酸で構成される遺伝子を用いて行われる。地球生命の場合、4種類の塩基をD―リボース(またはD―デオキシリボース)という糖と結びついた、化合物ヌクレオシドが、リン酸と結合してヌクレオチドとなり、これが鎖状につながって構成される。
この各塩基には、「塩基対」という水素結合で結びつきやすい組み合わせがあり、核酸は、必ずこの塩基対に応じたもう1本の核酸と対をつくる。これがDNAである。
対になったDNAを引き離すと、それぞれの核酸は、周囲から塩基を集め、対の相手を作り、その結果同じDNAが2組出来上がる。これが生命の自己複製の基礎である。
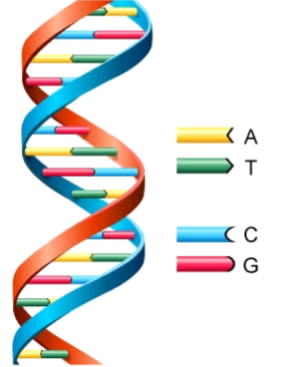
地球生命では、DNAの連なる塩基3つを1組とする意味を持ち、細胞を構成するたんぱく質のアミノ酸が、どのように並ぶかを、DNAから複製したm-RNAで規定し、親と同じ構造を作り出す。生命が自己複製を行うについて、地球外の生命でも、基本的に塩基対構造と似た働きを持つ物質を介すると考えられるが、地球環境内では、塩基以外に相応する物質はほとんど無い。
ただし、地球外生物では、使用する塩基の数が4種以外であったり、生体の基本物質を規定する塩基数は、3つ1組以外の組み合わせを利用する可能性も想定できる。
②エネルギー代(たい)謝(しや)
生命は、成長や増殖に必要なエネルギー源を、外部から栄養の形で得る。栄養はそのまま用いることができないため、複雑な化学反応をへてエネルギーに変換する。―これを代謝という。
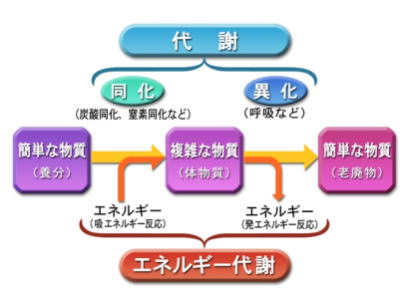
代謝は、生命の維持のために、有機体が行う外界から取り入れた無機物や有機化合物を素材として行う、一連の合成や化学反応のことで、新陳代謝の略称である。
これらの経路によって、有機体は、その成長と生殖を可能にし、その体系を維持している。代謝は大きく異化 と同化 の2つに区分される。
異化は、物質を分解することで、エネルギーを得る過程である。例えば細胞呼吸がある。同化は、エネルギーで物質を合成する過程であり、例えばタンパク質・核酸・多糖・脂質の合成がある。
③死
生物の細胞や臓器による生命活動が、不可逆的に失なわれることを死と呼ぶ。生命にとっては、その誕生とともに、その生命活動の終焉である、死も約束されているのである。
ただ、生命の定義が難しいのと同様に、死の定義も困難な問題である。そのため、生きている状態と死んでいる状態を、はっきりと区別することはできない。
多細胞生物では、個体の死と細胞の死は、別々に考えられるべきで、例えば、臓器移植の場合、臓器提供者が死亡しても、移植が成功すればその臓器は生きていると考えられる。
また生命体は普通、子をなしてその血統を存続させる。
これを細胞レベルで見れば、細胞の分裂と融合に基づく、連続性は常に維持されているため、その意味で生命は停止せずに連続していると表現する事も出来る。これを生命の連続性という。
多くの宗教では、何らかの形での「死後の世界」や輪廻、転生などが存在していると考えられている。
④進化
生命の進化とは、生物個体群の性質が、世代を経るにつれて変化する現象である。
また、その背景にある遺伝的変化を重視し、個体群内の遺伝子頻度の変化として、定義されることもある。この定義で、成長や変態のような、個体の発生上の変化は進化に含まれない。また狭義に、種以上のレベルでの変化のみを、進化とみなすこともあるが、一般的ではない。逆に、文化的伝達による、累積的変化や生物群集の変化をも、広く進化と呼ぶこともある。
進化の過程で、ある器官が単純化したり、縮小したりすることを退化というが、これもあくまで進化の1つである。退化は進化の対義語ではない。
生物は不変のものではなく、共通祖先から、長大な年月の間に、次第に変化して現生の複雑で多様な生物が生じたということが、膨大な証拠から分かっている。
進化論は、チャールズ・ダーウィンなど複数の博物学者が、動物や植物の分類学的な洞察から導きだした仮説から始まった。
現在の自然科学では、この説を裏付ける証拠が、形態学、遺伝学、比較発生学、分子生物学など、さまざまな分野から提出されており、進化はほぼ確実に起こってきた事実である、と生物学者・科学者からは認められている。
#目 次
極限環境での生命誕生
先に生命の定義で最小の単位が「細胞」であると定義したが、実は、生物発祥は、まだ細胞を有しない原始的微生物まで遡る。
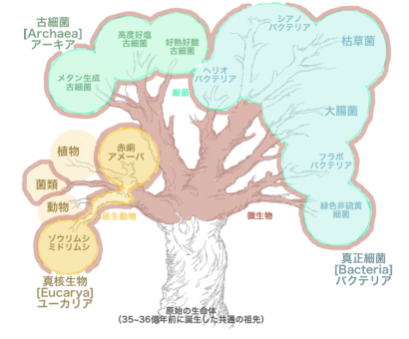
生命誕生のスタート地点となるのは、真正細菌と古細菌の既知のタンパク質遺伝子である。
最新の解析機で、数千もの微生物の遺伝子配列解析で、600万ほどの遺伝子がDNAデータバンクに蓄積された。ヒトとマウスで同じ機能を持つ遺伝子は、遺伝的には最初の哺乳類の先祖遺伝子の、共通の子孫という関係にある。
つまりDNAの配列を比較することで、進化の系統図を描くことができる。
600万遺伝子の解析の結果、真正細菌と古細菌の共通の祖先であるLUCA(単細胞有機体)が、355個発見された。遺伝子は、有機体が生きる環境に適応する。そのためLUCAが持っていた遺伝子を特定することで、それが生きた環境や生きた方法を知ることができる。
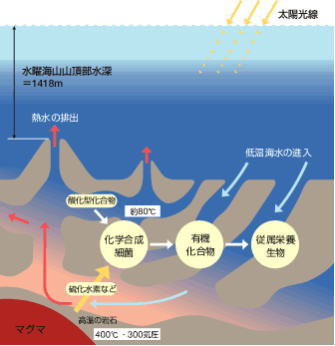
355個の遺伝子が、示していたのものは、「深海熱水噴出孔」の環境に住んでいた生物であった。深海熱水噴出孔では、ガスが充満し、金属が存在し、海底から噴出したマグマに触れた海水が、激しいホットプルームを発生させるという、生物にとっては極限環境の場所である。深海熱水噴出孔は、珍しい生命体が発見されていて、極端な化学的性質ゆえに、生命が誕生した場所ではないか。 ―と推測されてきた場所でもある。
LUCA由来の355個の遺伝子は、エネルギー源として「水素を代謝」するものと、酵素のための遺伝子を1つ含んでいる。これは極端に高温な環境で生きる微生物にしか見られないものである。 #目 次
極限環境に生きる古細菌
古細菌は、細菌の名を持つが、現在の生物分類上では、独立したドメインまたは界が与えられている。 一般には、メタン菌・高度好塩菌・好熱好酸菌・超好熱菌など、極限環境に生息する生物として認知されている。
形態的には、細菌と同じ原核生物(核を持たない細胞から成る生物)に属し、細胞の大きさ、細胞核を持たないことなどの点で共通する。
このため真正細菌と同じグループ(モネラ界)として扱われて来た。しかし分子レベルでの研究が進むにつれ、その違いが明らかになってきている。

古細菌
たとえば、古細菌とその他の生物、特に真正細菌の間には、さまざまな違いが知られている。さらに進化系統的には、むしろ真正細菌よりも真核生物に近縁で、DNAの複製やタンパク質合成系といった、生命の基幹部分の機構が真核生物に類似している。このような生化学的差異、系統学的位置が明らかになるに従い、独立のドメインとして扱われるようになった。
その生体システムは、未だ不明な点が多いが、原始生命体や真核生物の起源、あるいは有用酵素の利用・メタン発酵などと関連して研究が進められている。
生息環境は、生物圏の広い範囲に分布し、最大で地球上の総バイオマス(生物体量)の20%を占めるとも言われている。
純粋培養の可能な古細菌の多くは、極限環境微生物、あるいは非常に強い嫌気度を要求するメタン菌であり、このため歴史的に、極端な環境に分布すると考えられてきた。
好熱菌
現に、100℃を超える高温で活動する古細菌が、間欠泉やブラックスモーカー、油田から発見された。また、高い塩濃度や強酸、強アルカリ、非常に低温の環境からも、比較的容易に古細菌を発見することができる。
一般に、極限環境に生息する古細菌は、3つの生理的なグループ、高度好塩菌、超好熱菌、好熱好酸菌へと区分することができる。
好塩性の古細菌― ハロバクテリウム属を含む高度好塩菌は、20~25%の塩化ナトリュウム濃度で盛んに増殖し、塩湖など非常に塩濃度の高い環境に生息する。一方、好熱菌は、温泉など45℃以上の環境で 高度好塩菌 よく活動する。このうち80℃以上の生育温度を持つものを超好熱菌と呼ぶが、メタノピュルス・カンドレリ116は、全生物中最も高温で生育する生物として知られ、122℃で増殖が可能と報告された。
高度好塩菌
また強酸を好む好熱好酸菌は、スルフォロブス目や、テルモプラズマ綱に代表され、温泉や硫気孔、ボタ山などから発見される。アルカリ性の塩湖には、高度好塩好アルカリ菌が生息する。古細菌に有性生殖はなく、真正細菌と同様、単純な二分裂によって増殖(繁殖)する。 出芽により増殖するテルモプロテウス目など一部を除くと、分裂後も殆ど同じクローンが2体できるだけである。最適条件での増殖速度は約30分、遅いものだと数日を要する。分裂に伴う細胞膜の切断や、DNAの分配は、真正細菌に似ていると言われている。#目 次
人と共生する真正細菌
真正細菌とは、いわゆる細菌・バクテリアのことで、大腸菌、枯草菌、シアノバクテリアなどを含む生物群である。形状は、球菌か桿菌、ラセン菌が一般的で、通常1~10
μmほどの微小な生物である。
大腸菌
核を持たないという点で、古細菌と類似するが、古細菌―真核生物にいたる系統とは、異なる系統に属し、両者はおおよそ35~41億年前に分岐したと考えられている。
遺伝やタンパク質合成系の一部に、異なる機構を採用し、ペプチドグリカンより成る細胞壁、エステル型脂質より構成される細胞膜の存在で、古細菌とは区別される。
真正細菌は、地球上のあらゆる環境に存在しており、その代謝系は非常に多様である。
個体数は5×1030と推定されており、その生物量も膨大である。
光合成や窒素固定、有機物の分解過程など、物質循環で非常に重要な位置を占めている。食品関係では、チーズ、納豆、ヨーグルトといった発酵過程で、微生物学発展以前から用いられてきた。
また、腸内細菌群は、食物の消化過程には、欠かすことのできない要素である。
一部のものは病原細菌として、ヒトや動物の感染症の原因になる。
対立遺伝子を持たず、遺伝子型がそのまま表現型をとり、世代時間が短く変異体が得られやすい。あるいは形質転換系の確立などもあり、近年の分子生物学を中心とした生物学は、真正細菌を中心に発展してきた。
これまでストレプトマイシンや、クロラムフェニコール、テトラサイクリンなどなど様々な抗生物質が発見されてきた。またその製造や免疫系の新薬開発の上でも、非常に重要である。#目 次
全ての生物 ―真核生物
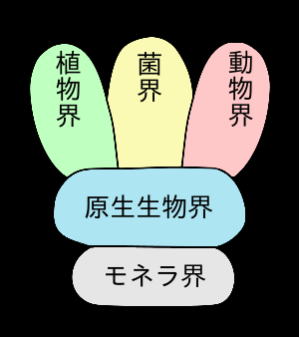
真核生物とは、生物の二大群の一で、細菌類と藍藻類を除く大多数の生物を含む。
つまり、動物界、植物界および原生生物界を構成する、全ての生物をいう。 原生生物界には原生動物(原虫)、真菌、単細胞藻、粘菌(変形菌)が属している。
原生生物からヒトを含む哺乳類までの細胞は、全て真核細胞であり、基本的に細胞構造や機能は同じ。
真核細胞は、中心に核膜で包まれた核をもち、その中に複数個の染色体が存在する。染色体はヒストンなどの塩基性タンパク質と結合した二重らせん(2本鎖)DNAである。核内には核小体(仁)もみられることがある。
一般に有糸分裂で増殖するが、藻類、原虫、真菌、粘菌のように、胞子または鞭毛をもった遊走子や、栄養細胞自体の発芽で増殖する場合もある。
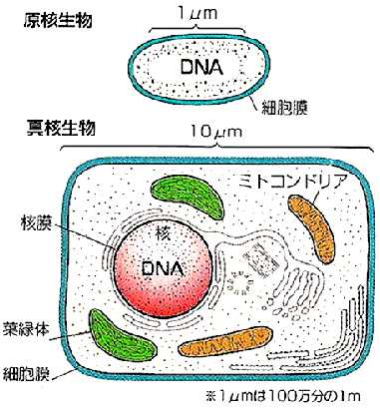
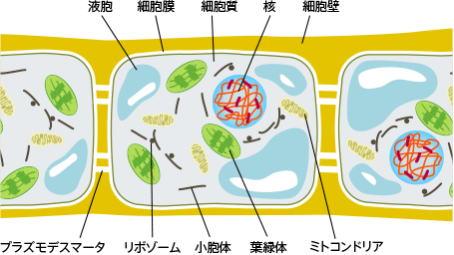
細胞質内には、複雑に分化した膜系の小器官が存在する。
呼吸代謝を行うミトコンドリア、光合成を行う葉緑体、タンパク質を合成するリボゾームが、多数存在する小胞体、種々の加水分解酵素を含んで高分子物質を消化するリソゾーム、ムコ多糖などを合成するゴルジ体などは重要な膜系の小器官である。
細胞質は薄い細胞質膜で包まれているが、高等植物、単細胞藻類および真菌の細胞には細胞質膜の外側に厚い細胞壁がある。しかし、高等動物や原生動物の細胞には細胞壁はない。
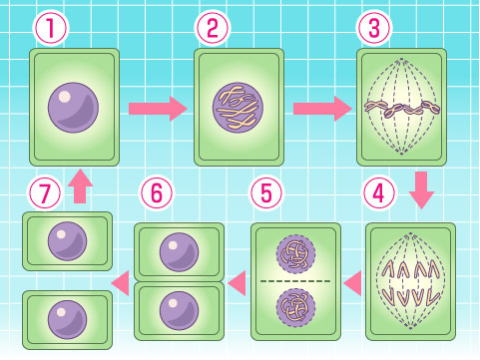
細胞分裂には、まず核分裂が行われる。一般に、核内のDNAは細胞分裂に先立って、より集まった染色体になり、DNAはこの染色体ごとに新しい2つの細胞のために糸状の構造(紡錘糸)によって分かれ、分かれた染色体のコピーが渡される。これを有糸分裂と呼ぶ。
真核生物の染色体は直鎖状であり、末端にはテロメアと呼ばれる構造がある。ほとんどの真核生物では、有性生殖が行われる。
減数分裂後、染色体の半数体を2つ合わせて核の合成を行う。これには様々なパターンが存在する。 #目 次
多細胞生物ま誕生
多細胞生物は、複数の細胞で構成されている生物のこと。
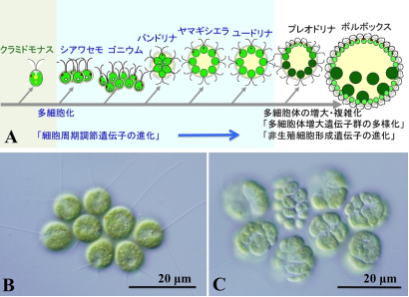
核を持ち細胞分裂のときに、染色体構造を生じる真核生物は、その誕生から数億年は、単細胞生物として存在していたが、14~10億年前に、多細胞生物が誕生した。
最初は、単なる細胞の集合体であったが、やがて細胞がそれぞれ特別の機能を果たすようになった。
生物の最大の課題は種の保存である。生物の生も死も、種を保存する為に存在している。そして種を保存していくには生殖、さらに生存し続けいていくには、栄養摂取が最大の課題となる。生殖と栄養摂取の2つの課題を、バランスよくこなしていくことが、生物に求められる。
つまり単細胞から、多細胞への進化とは、単純化すれば保存細胞と仕事細胞の分化と見てよい。生殖というのは生物にとって最大の課題であり、最大の負担でもある。単細胞から、多細胞へ変化していく過程は、体細胞を担う仕事細胞と、生殖器官を担う保存細胞に、役割分化したことが始まりである。
多細胞を司る細胞群は、単細胞に戻れないという細胞の不可逆性がある。
そして、多細胞生物の中で、唯一の単細胞である精子や卵子は、それ単独では存在できない。つまり精子も卵子も合体する事で、初めて生存が可能になったのである。
単細胞では生き残れない、厳しい環境を克服したのが多細胞生物システムである。
その結果、体細胞はますます進化したが、ところが「生殖細胞」だけは、下等動物から高等動物まで殆ど進化していない。
つまり保存細胞を犠牲にすることで、進化を遂げたのが多細胞生物なのである。その頂点に立ったのが、動物群である。
多細胞生物の出現は、生物の複雑化・大型化への道を開き、9億年前に始まった有性生殖へと変化し、やがて多彩な生物へと進化していく。
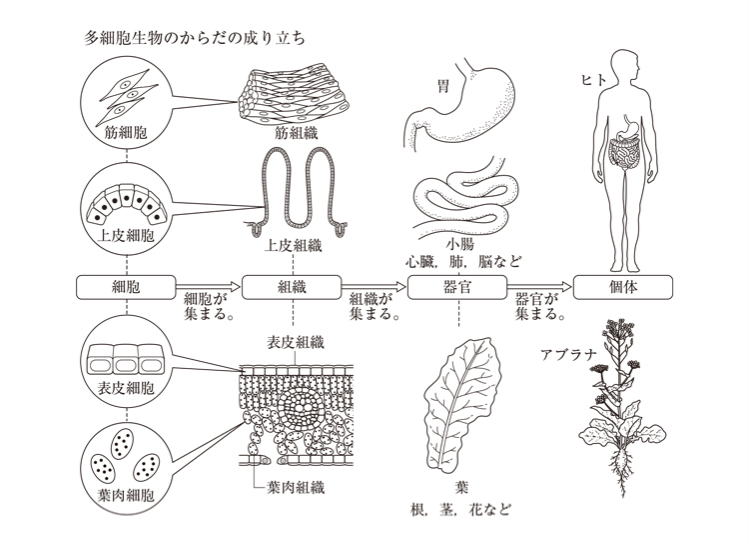
動物界や植物界に所属するものは、すべて多細胞生物である。
菌界のものには、多細胞生物と、若干の単細胞生物が含まれている。肉眼で確認できる大部分の生物は、多細胞生物である。
細かく見れば、原核生物にも、簡単な多細胞構造を持つものがあり、真核の単細胞生物が多い原生生物界にも、ある程度発達した多細胞体制を持つものが含まれる。
多細胞体制の進化は、その分類群により様々な形を取る。
その生物の生活と深く関わりがあるので、菌類的なもの・植物的なもの・動物的なもの、それぞれに特徴的な発達が見られる。
最も少ない細胞数で構成されている生物は、淡水生の藻であるシアワセモの4個である。アオミドロやコンブなどは、同じ形の細胞で出来ているが、複雑な生物では、体の部分によって、細胞の形や働きが違っている。
形も働きも同じ細胞が集まっているのが、組織がいくつか集合して1つの働きをなす器官をつくりあげている。生物の体は、複雑なほど、組織や器官が複雑に結びついている。動物と植物とでは、組織や器官の造りや働きも違っている。 #目 次
植物の多細胞体制
地球上の生物は菌類、植物、動物に大別されるが、植物とは、細胞壁を有し、独立栄養で光合成を行うことができる生物をいう。菌類も細胞壁をもつが、もっぱら動植物に寄生し、生活のすべてをこれらの生物に依存している。
動物は細胞壁をもたず、栄養源は植物に依存している。このように、植物は生物の栄養源として重要な役割を果たしている。
こうした背景で、植物は、細胞膜の外に細胞がセルロースなどの細胞壁で覆われている。その細胞内には葉緑素を含み、光合成を行い、無機物から有機物を合成する。
また発生の際に、一次生長を行う部分が、軸の両端部に限られ、しかもその生長が連続的、かつ無限の能力をもっている。
細胞壁は、組成の簡単な植物体の保護の役割があり、乾燥から守り、とくに陸上植物では植物体を支えている。陸上植物の細胞壁は、おもに多糖類のセルロースで、藻類では、マンナン、グルカン、アルギニン酸、寒天質などの細胞壁をもち、セルロースを欠くものもある。
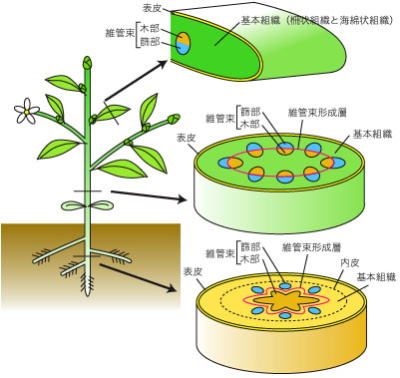
光合成をするためには、光が当たって、二酸化炭素と水の取り込みができれば、他に何もいらない。体を固定する必要などから、多細胞化をするにせよ、細胞が平らに並んでいる構造までは、特に工夫しなくても各細胞が自活できる。
実際、いくつもの藻類では、独自に多細胞化が進み、原核生物の藍藻にも、一列に並んだ細胞からなる、多細胞体制がある。
その多くは、生殖細胞の分化以外に、はっきりとした分化は見られず、生殖細胞が分化していないものも多い。
植物の組織分化は、陸上進出時に大きく変化した。体を支え、水を吸い上げるための根や維管束、水の少ないところでの有性生殖を確保するための花など、水中生活の藻類とは遙かに異なった、段階の多細胞体制の進化が起きている。 #目 次
動物の多細胞体制
動物的活動は、移動をしながら、外部から食料を摂取する。
このような方法の生物で、多細胞のものは、動物界に属するものだけであり、原生生物界の動物的生き物はすべて単細胞、ないしは群体である。 この方向での多細胞化の進化が、どのように行われたかについては、今も諸説がある。
決定的な定説はないが、有力な説として、ヘッケルの「系統発生論」がある。この論は、生物の種族の発生から、絶滅するまでの形態変化の系統のことである。
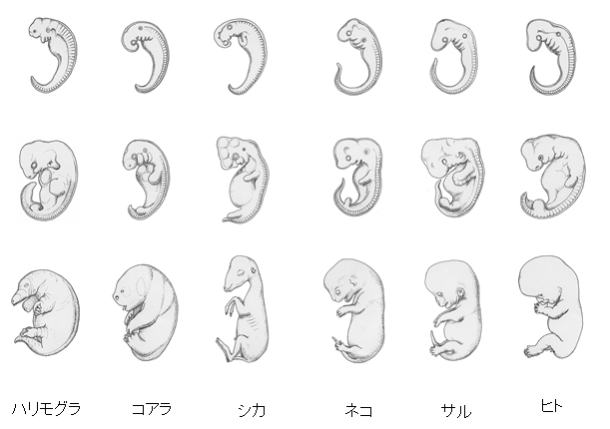
ヘッケルの「系統発生論」
ある生物種族が、進化とともに形態を変え、一つの発展系統をつくりだすこと。
換言すれば、種族の進化の歴史ということになる。
ヘッケルは「個体発生は、系統発生を繰返す」という表現で、両者の関係を端的に表わした。 ただこれは簡略にすぎる表現であり、いろいろな批判を受け、一時は否定された。今日では、個体発生と系統発生の関係を表わす一形式として、他の形式とともに認められている。
いずれにせよ、移動することで、進行方向の細胞が、より餌をとる可能性が高いから、早い段階で、栄養が各細胞に行き渡る仕組みが発達したと思われる。
同様に、運動の仕組み、感覚、及びそれらの情報を伝える仕組みが発達し、複雑な組織系、器官系を進化させた。その代償として、個々の細胞は互いに依存しあい、個々の部分が独立して生き延びる能力は低くなっている。
なお、変形菌も、ある意味で動物的活動である。これらの場合、多細胞化ではなく、核が増えても細胞分裂をせず、多核体化することを選択したとも見られる。
#目 次
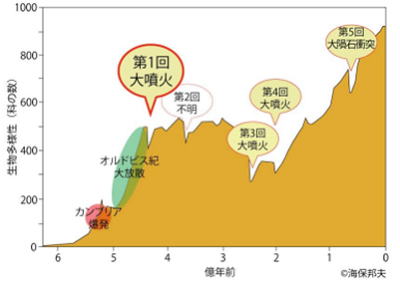
カンブリア爆発
極端な寒冷期であったマリノアン氷河時代は、約6億3千500万年前に終了した。
カンブリア紀は、約5億4千200万年前に始まり、約5億年前までの期間で、その気候は、地球の気候が温暖化しつつある時期に相当する。カンブリア紀の特徴は、動物の多様性が一気に増加したことである。

カンブリア紀以前に確認されている動物の門は、「刺胞動物」(クラゲ型は鐘状、ポリプ型は円筒形をしている)と「海綿動物」(外形は円筒状・樹枝状または不規則な団塊状で、体の下面で岩礁その他のものに付着する)で、カンブリア紀の代表的地層の、カナダのロッキー山脈のなかにあるバージェス頁(けつ)岩(がん)からは、この2門以外に節足動物、腕足動物、脊索動物、軟体動物、棘皮動物、環形動物、袋形動物、脊索動物などが確認されている。
ついでながら、頁岩というのは、海底で泥(16分の1㎜以下の細かな粒子)が、水平に堆積してできた岩石の中で、頁のように薄くはがれる性質をもつ岩のことである。
このように突然、大量の生物種が発生したことから、この事件は「カンブリア爆発」と呼ばれている。 これらの生物の多くが、リン酸塩やカルシウム塩からなる、固い殻を有していたため比較的化石として残りやすく、このことが顕生代開始の定義につながった。
生物が固い殻を持った理由として、カンブリア紀初期に、大量のリン酸塩が浅海に供給され、生物がリン酸塩を利用しやすかったこと、前時代には存在しなかった、強力な捕食者(アノマロカリスやオパビニアなど)に対抗する防御が必要になったと、推定されている。さらに捕食者から身を守るために、優れた遊泳能力や海底の泥に深くもぐる能力などが発達した。
これらの必然性から、生物の多様化が進んだと考えられている。
カンブリア紀の浅海における生物量としては、節足動物の三葉虫と腕足類が優勢であった。カンブリア紀に発生した生物の中には、現在見られない形の動物も多く存在したが、それらの動物は、カンブリア紀末までに大部分が淘汰された。 「三葉虫」の硬
骨格や腕足類の殻は、化石として残りやすいため、古くから世界各地で産出しており、1970年代まで「カンブリア紀」の生物相は、このような硬骨格生物が主体であると考えられてきた。

捕食者アノマロカリス
カンブリア紀の捕食者として有名な、「アノマロカリス」の全体像が解明されたのは70年代で、上記「バージェス頁岩」の研究による。その後80年代からグリーンランドのシリウスパセットや、中国の澄江(チェンジャン)での化石発掘と研究により、カンブリア紀の多様な生命の状況が判明した。
#目 次
カンブリア爆発の原因
カンブリア大爆発は、カンブリア初期に一斉に生物の体制が出そろった現象をいう。
これはスティーヴン・ジェイ・グールドが提唱した説で、彼はは、カンブリア紀に生物の体制の種類が爆発的に増加し、その後は減少に向かっていると主張した。
これは偶然か、自然選択では説明できない何らかのメカニズムが存在することになる。
その後の分子遺伝学の進歩から、実は、遺伝子の爆発的多様化は、カンブリア爆発のおよそ3億年前に起こっていることが分かった。 このため、カンブリア初期に、短期間に大進化が起こったわけではないとの考え方が主流となった。
つまり、カンブリア爆発は、「化石記録の」爆発的多様化であり、必ずしも進化的な爆発を意味しない。
リチャード・ドーキンスは、カンブリア紀あるいはそれ以前に、特殊な総合説では説明できないような進化現象が起き、生物の体制が出そろったというグールド以来の視点、爆発という概念自体に批判的である。

彼に依れば、現代の脊椎動物と、無脊椎動物が根本的に異なっているのは、両者が長い地質学的時間の間に、異なる方向に進化してきたからである。
少なくとも現在の証拠からは、種分化した当初から、全く異なる体制を持っていたと考える理由はない。 ―と主張する。
1998年、進化生物学者で古生物学者のアンドリュー・パーカーは、カンブリア爆発の原因として、有眼生物の誕生による、淘汰圧の高まりをあげた「光スイッチ説」を提唱した。
生物の歴史上、はじめて眼を持った生物(三葉虫)が生まれ、積極的に他者を捕食することによって、眼を持たない生物に対して有利となった。眼と、硬組織を獲得した生物が、その捕食に対抗できるようになったという説である。
そのために化石記録は、短期間で爆発的に多様化したように見える。パーカーは、カンブリア爆発を「多くの門が、同時期に一斉に硬組織を獲得した現象」と推定している。カンブリア爆発の原因として、スノーボールアース(雪球地球)の終結との関連性が、従来から指摘されていた。しかしパーカーは、スノーボールアース終結からカンブリア爆発まで、少なくとも3千200万年も経過しているから、関係は間接的なもと述べている。

約10億年前に、多細胞生物が出現し、その後、8億~6億年前に、スノーボールアースの間でも、生物は存在し続けた。
多細胞生物は原口を獲得し、強力な捕食能を有するに至った。 海底には、熱水鉱床などの、熱水を発する箇所があり、スノーボールアースの間、その近辺で生物は隔離されて生存したと考えられる。このような地理的な隔離は、ガラパゴス諸島やオーストラリア大陸のように、生物の多様性を形成する。
スノーボールアースの地理的な隔離の間、どのように捕食するか、どのように捕食から逃れるかの観点から、多細胞生物は多様性を形成し、これがエディアカラ生物群や、バージェス動物群のような多様性を形成し、スノーボールアース終結からカンブリア爆発まで、少なくとも3千200万年も経過していることから、その間、全地球的な捕食と被捕食の生存競争が存在したと考えられる。
バージェス動物群に見られる、アノマロカリスやオパビニアなどの大型捕食動物の出現とともに、カンブリア爆発の際には、堅い外骨格をまとった動物が、多く見られるようになった。
バージェス動物群とは、バージェス頁岩の中から、化石として発見された動物群である。いわゆるカンブリア爆発の後に発生した、多様な生物群が含まれることで有名。
エディアカラ生物群は、新たに出現した捕食動物に食い尽くされ、絶滅したとも言われている。#目 次
ビッグファイブ ―生物大量絶滅
地球上の生物は、その原初から数えて過去に5回、大規模な環境変動に伴う大絶滅を経験しているとされている。これをビッグファイブと言う。
大量絶滅は、地質時代区分で幾度か見られる現象である。
そもそも地質時代の「代」や「紀」の区分は、化石として発見される動物相の相違によるもの。
そもそも原生代・古生代・中生代・新生代の「代」の時代区分は、大量絶滅により従来の動物の多くが絶滅し、新たな動物が発生したことによる区分である。
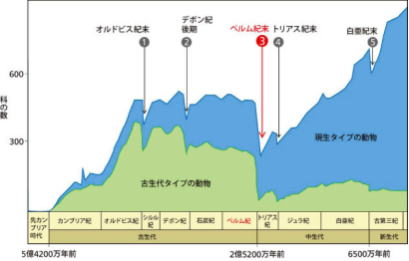
「紀」の時代区分は、「代」との比較では、動物相の相違は小さいが、大量絶滅による場合もある。多細胞生物が現れたエディアカラン以降、5度の大量絶滅 ―それぞれ、オルドビス紀末(O-S境界)、デボン紀末(F-F境界)、ペルム紀末(P-T境界)、三畳紀末(T-J境界)、白亜紀末(K-Pg境界)の大絶滅のことである。
この他に、若干規模の小さい絶滅が数度あったとされる。
生物大量絶滅の原因
大量絶滅の原因については、K-Pg境界のように、隕石や彗星などの天体の衝突説が有力視されている事件や、P-T境界のように、超大陸の形成と分裂に際する大規模な火山活動による環境変化)が有力視されている。
このようにさまざまであり、その原因は一定しているわけではない。大量絶滅の直後には、空席になったニッチ(生態的地位)を埋めるべく、生き延びた生物による、急激な適応放散がおきる。 例えば恐竜が絶滅したことにより、白亜紀以前には小型動物が中心であった哺乳類[は、急速に多様化・大型化が進み、生態系の上位の存在として繁栄を享受することになる。

アリゾナにある隕石クレータ
大量絶滅の最後のもの、白亜紀末の大絶滅は、恐竜の絶滅で有名だが、これは最近では「隕石衝突が原因」と ー いうのがほぼ通説になりつつある。
なお、これらの大量絶滅クラスの火山の大噴火というのは、火山が火口からマグマを噴出させると言った次元のレベルではない。一説によると、スーパーホットプルームと呼ばれる、地球のコアの熱によって暖められた大きな塊が、地表に丸ごと噴出し、凄まじい熱反応を引き起こしたのが、これらの大絶滅の原因だったのではないかと言われている。 #目 次
オルドビス紀末の大量絶滅
オルドビス紀は、顕生代の5大量絶滅事変の1つに数えられる大量絶滅により、その終焉を迎えた。このオルドビス紀末の絶滅事変は、絶滅した属の割合で見ればペルム紀末の大量絶滅に次ぐ大規模なものである。海生多細胞生物の科の22%、属の49%%が絶滅し、腕足類、コケムシ類も大打撃を受けた。
大量絶滅の原因は、はっきりしていないが、大規模な火山活動によって、寒冷期を迎えたのではと推測されている。

この時期、大陸は南極域にあり、短い期間ながら大陸氷河が発達した。
絶滅は、氷床の発達に伴う海水準の低下時、及び氷河の消滅に伴う海水準の上昇時の、2回確認されている。オルドビス紀末の大噴火により、噴出した塊は水銀を含んでいる。これが噴火活動によって、空高く吹き上げられ、地表に水銀が堆積する。今回の発表は、この痕跡が見つかったことによって、オルドビス紀末の大絶滅が火山活動由来であると証明されたことによるものである。
ちなみに、このクラスの大噴火は、成層圏に到達した硫黄が硫酸になり、太陽光を反射して地球全体の寒冷化を招き、多くの生物を死に追いやるという。オルドビス紀末の大絶滅は、当時存在した種の85%が死に絶えたとも言われている。
オルドビス紀末の大絶滅が、火山活動を原因とするものであったことが突き止められた。オルドビス紀末大量絶滅を記録した、2カ所の地層から、水銀の濃集が発見された。5度の生物の大量絶滅のうち、後半の3つの大量絶滅の原因はほぼ明らかだが、最初の2つの大量絶滅は、未解明のままとなっていた。

4億4千500万年前~4億4千300万年前の、最初の大量絶滅では、カンブリア爆発とオルドビス紀大放散を経て、形成された珊瑚礁と頭足類、海サソリ、筆石類、三葉虫、腹足類など、多様な動物群が大きな影響を受け、科の5分の1、属の半数が絶滅した。
その大量絶滅を記録する中国とアメリカの地層から、堆積岩試料を採取し、元素分析を実施し、水銀の濃集が発見された。
水銀の濃集は、異なる時代の3回の大火山噴火時にも認められており、今回発見された濃集も大火山噴火で、マントル中に起源を持つ水銀が、空高く放出され世界中に堆積したものと考えられる。
大火山噴火では、成層圏に放出された大量の二酸化硫黄ガスが、硫酸となり地球を覆い、太陽光を反射することで、地球規模の寒冷化が起きる。
また、この時期の前後の地球は、氷床がない温暖期で、この時期のみ氷床があったことから、それがこの大量絶滅に関係していると考えられていた。したがって今回の発見は、大規模火山噴火が寒冷化を招き、氷床発達と大量絶滅を引き起こしたことを示唆している。今回の成果から、5度の生物の大量絶滅のうち、3度は大規模火山噴火によるもの、1度は小惑星衝突によるものである。
#目 次
デボン紀後期(F-F境界)の大量絶滅
今から3億7200万年前、デボン紀末に80%ほどの生物種が絶滅した。ところが、大量絶滅を引き起こす程の、極限的な自然環境変化は確認されていない。
デボン紀にも隕石の衝突があり、スウェーデンに直径は50㎞の「シャン・クレーター」が残っている。
しかし白亜紀末の生物大量絶滅を引き起こした、巨大隕石の衝突跡であるユカタン半島のチチュルブ・クレーターは直径160㎞もある。これに比較すると、デボン紀の隕石は、比較的小さく、生物大量絶滅に至るほどの破壊力はなかったと思われる。
デボン紀の大陸と海
一方、デボン紀の生物大量絶滅には、ひとつの特徴がある。
デボン紀の時代は「魚の時代」といわれ、魚類が大繁栄し、サメの登場やイクチオステガなどの陸生の脊椎動物が現れるなど、大躍進の時代である。
魚類の「板皮類」や「棘魚類」では、海に住む種類と、湖や川などの淡水域に住む種類で、絶滅率に明確な差があった。
板皮類では海に住む種の65%が絶滅したのにに対し、淡水に住む種は23%程度であった。また海に住む棘魚類は特に深刻で、87%も絶滅しているが、淡水に住む種は30%程度にとどまっている。いずれも海生種に深刻なダメージを与えているという特徴が有る。

魚類に限らず、デボン紀には、節足動物など陸の生き物より、三葉虫や、腕足類、サンゴなど、海の生き物の絶滅が顕著である。
つまり、デボン紀の生物大量絶滅は、海の中で主に起こったことがひとつの特徴である。またう一つ、腕足類という二枚貝に似た生物では、赤道近くの低緯度の熱帯の海に住む種類は91%と壊滅的であったが、高緯度の冷たい海に住む種類は27%にとどまっている。
この絶滅率の差から、大規模な寒冷化が起こったものと考えられる。海水温が下がるとすると、高緯度に生息していた腕足類は、低緯度の海域に移動すれば、生き長らえるが、低緯度の熱帯の海に棲んでいた腕足類は逃げ場がなく、多くが絶滅したと考えられている。
しかし、デボン紀の生物が大量に絶滅した時期に、現在の南極のような大規模な寒冷化によってできる、発達した氷床の痕跡が見当たらず、デボン紀の生物大量絶滅の原因については、まだ確実にいえることはない。#目 次
超酸素欠乏―スーパーアノキシア
スーパーアノキシア(超酸素欠乏事件)とは、P―T境界で起こった大規模な海洋無酸素事変である。
世界各所に産出する、当時の海洋起源の堆積岩(泥岩やチャートなど)の研究から、約2億5千100万年前の、前後約2千万年にわたって、海洋が酸素欠乏状態にあったことが判明している。
地球史上では約百万年程度の、酸素欠乏事件は何回か発生しているが、全海洋規模かつ約2千万年という長期間にわたる酸素欠乏が起こったのは、P―T境界のみである。
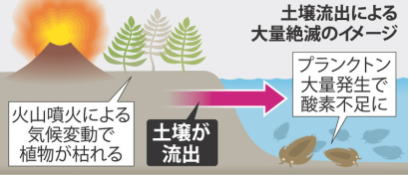
スーパーアノキシアは、P―T境界の前段階の、ガダルピアン末の大絶滅と同じ時期の、2億6千万年前に始まり、最盛期はP―T境界に一致している。最盛期には、その前後の地層にふんだんに見られる放散虫の化石が、全く消滅しており、大洋の表層でも大量絶滅が起こっていたと考えられる。P―T境界における酸素欠乏については、「大絶滅により、光合成を行う生物が、極度に減少した結果、海洋中の酸素が減少した」という考え方と、「何らかの原因で海洋が低酸素化した結果、呼吸できなくなった生物が、大量に死滅した」という二通りの解釈がなされている。#目 次
ペルム紀末(P-T境界)の大量絶滅
ペルム紀の末の約2億5千万年前に、P―T境界で、史上最大規模の生物大量絶滅事件が起こった。古生代最後のペルム紀 (Permian) と、中生代最初の三畳紀
(Triassic) の境目であることから、両者の頭文字を取って「P―T境界」と命名されている。

地質学では、時代境界を規定するため、新しい生物種の出現の時期を利用している。このため、地質学上の時代境界の地層と、生物の大量絶滅時期の地層とは一致していない。つまり、大絶滅の時期を、地質学に基づいて厳密に規定すると、「後期ペルム紀」が、大量絶滅となる。
一般に、古生代の陸上生物は、両生類や単弓類、中生代は、恐竜に代表される爬虫類の時代と言われる。P―T境界では、この交代の原因となった、大量絶滅事件が起こった。
ペルム紀末に、海中に住んでいた海生無脊椎動物は、種レベルでの絶滅率は90%以上、82%の属、半分の科が消滅したと見積もられている。この中には三葉虫・古生代型サンゴ・フズリナなど、古生代に幅広く生息していた生物種が含まれる。
脊椎動物では、82%の科が絶滅した。また昆虫・植物などの陸上生物も、多くの種類が絶滅した。絶滅した生物種は、恐竜の絶滅で有名なK―T境界よりはるかに多く、カンブリア紀以降で最大規模の絶滅であった。
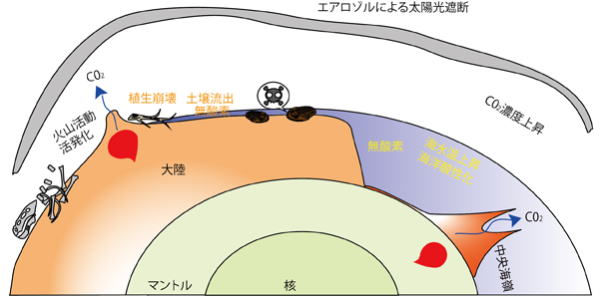
| 巨大火山活動が成層圏エアロゾルを形成、気候変動を起こし、陸上植生の崩壊と土壌流出事件を起し、浅海無酸素事件により、1段階目の海の絶滅、その後、火山活動による大気温室効果ガスの増加により、極端温暖化、海洋酸性化、海洋無酸素状態のピークにより2段階目の海の絶滅がおきたとれさる |
大絶滅の原因については、種々の仮説があるが、いまのところ地質学者の大半が同意する明確な説は無い。地質時代の年代分析では、90年代以降、新しい分析技術に基づいた研究が著しく進んだ。
このP―T境界について、それまでは何百万年も続いたと考えられてきたが、1994年、スタンリーとヤングが、ペルム紀末の絶滅が、800万年から1千万年の間を隔てた、2回の大量絶滅であること発表した。さらに1996年、アメリカのノルが、「絶滅事件は、約2億6千万年前と、約2億5千万年前の2回起こった」と発表した。
最初の2億6千万年前の事件は、ペルム紀中期に相当し、ガダルピアン世の末期に相当する。
この絶滅事件は、海水準が突然低下し、多数の海洋生物が絶滅したとされており、陸上生物についても、環境変化による大量絶滅があったとされている。

2番目の事件が、古生代の生態系が壊滅した、破局的な大量絶滅に相当する。2番目の(本来の)大絶滅事件は、中国南部の煤山にある、当時の礁(しよう)の地層に挟み込まれた、複数の火山灰の分析から、2億5千160万年前に、突然絶滅が始まり、つづく百万年で、大絶滅が起こったと想定されている。
この年代値は中国の煤山と、そこから千㎞離れた中国広西壮族自治区にあるP―T境界層で、同じ値が得られている。
他の研究では、この絶滅イベントは、ペルム紀末と、三畳紀最前期 の2回に分かれ、前者では大半の浮遊生と、若干の底生生物を中心に56・5%の種が絶滅し、後者では、残った内の70・9%種が絶滅したと結論づけている。海洋生物の多様性の推移について、海洋生物の科が、化石として最初に現れてから、最後に消えるまでを記した一覧表が作製された。
その結果、約2億5千万年前に、もっとも大きな大量絶滅が起こっていることが判明した。研究者は海洋生物の進化度を、カンブリア型進化動物相、古生代型進化動物相、現代型進化動物相の3種に分類し、各々の科の消長を記録した。
カンブリア型進化動物相は、三葉虫に代表される最も古い動物相で、ペルム紀前の石炭紀に、すでに大幅に衰退していたが、ペルム紀末にすべて絶滅した。
古生代型進化動物相は、ペルム紀当時の浅い海で最も繁栄していた相で、有関節腕足類、古生代型サンゴ、アンモナイト、ウミユリ、レース状コケムシ等で、P―T境界で72%の科が絶滅した。現代型進化動物相は、二枚貝、腹足類、甲殻類、硬骨魚類、軟骨魚類で、科レベルでの絶滅率は27%であった。
#目 次
陸上の動物と植物の絶滅
陸上の動物にも、ペルム紀末に2段階の大量絶滅が起こった。爬虫類では、ペルム紀中期に生息していた26属のうち、ガダルピアン末に相当する、約2億6千万年前に9属が絶滅した。その後、多様性が回復し44属に回復したが、P―T境界後に、再度大きな影響を受け、生き残ったのは7属だけとなった。
単弓類(哺乳類及び、古くは哺乳類型爬虫類とも呼ばれた、その祖となる生物の総称)では、ディキノドンなどの単弓類は、P―T境界で35属が2属に減った。
昆虫類では、昆虫類の歴史でP―T境界は、唯一の大量絶滅事件である。ペルム紀に22目が生息していたが、そのうち8目が消滅し、他の5目が多くの科を失った。
昆虫類はその後は、1~2目しか絶滅していない。
植物では、古生代は沼沢林のシダ類、ヒカゲノカズラ類および木本シダが繁茂していたが、中生代には針葉樹、イチョウ、ソテツおよび現生シダに置き換わった。
古生代の石炭紀から続く、ペルム紀の地層には、石炭が大量に埋蔵されているが、P―T境界を境に、石炭が突然無くなる。
これは全世界的に同時に起こっており、南極、オーストラリア、インド、中国などで確認されている。 厚い石炭層が再度出現するのは、中期三畳紀の終わり、もしくは後期三畳紀からである。
石炭の消滅の原因は、石炭の元となる泥炭が生成する湿地帯が、長期にわたる気候変動(温暖化等)で、消滅したとも考えられる。
しかし全世界的に、一斉に石炭が消えていることから、気候変動だけではなく、湿地帯に生息していた植物類が、消滅したと考えられている。
またP―T境界を境に、平野部の河川の流れ方が、急激に変わったことが知られている。ペルム紀の湿潤な平野部の河川は、現在と同様に蛇行しながら、ゆっくり流れ、流域には、泥岩が多く堆積していた。しかしP―T境界を境に、突然、泥岩が減り、砂岩や礫岩が出現する。また河川の流れ方も、蛇行が減って、網状に流れるタイプに変った。 これはP―T境界において、流域の植物が壊滅的に減少し、土壌が無くなったこと、気候がより温暖・乾燥化したことによる、雨水による浸食が激化したためと考えられる。
この突然の変化は、南アフリカ、オーストラリア、ヨーロッパ、ウラル山脈の南部、インドなど、世界各地のP―T境界の地層で確認されている。#目 次
三畳紀末(T-J境界)の大量絶滅
三畳紀の終わりに、地球上の陸と海の生物の、少なくとも半分が絶滅した。
これを契機に、恐竜が勢力を拡大し、その後1億3千500万年にわたって、地球を支配することになる。この大量絶滅の原因は、大規模な火山噴火と考えられていたが、その正確な年代は特定できていなかった。
近年の研究では、多数の動植物種の化石が消える時期に、非常に大規模な溶岩の噴出が起こっていることが確認された。この溶岩が、アメリカ大陸に堆積したと仮定すると、その高さは90mを超える計算になる。

タイミル半島プトラナにある洪水玄武岩流
2億100万年ほど前、超大陸パンゲアが、プレート運動によって分裂を始めたが、このとき、地殻の下にあるマントルの岩石が溶け、これほど大きな噴出につながったという。このとき噴出した溶岩は、「洪水玄武岩」と呼ばれている。
60万年の間に4回に渡って噴出したが、そのうち最初の火山活動が、多くの生物種の絶滅につながったと見られている。
古代ワニの祖先、コノドント ―ウナギのような生物、哺乳類型爬虫類とも呼ばれる獣弓類など、多数の生物種がこのとき絶滅したとされている。つまり、この時期を境に、全てが化石として残らなくなっているからだ。
洪水玄武岩とは、大陸地域で、非常に膨大な量の玄武岩質溶岩が噴出し、その玄武岩が巨大な岩体として台地を形成した。その地形から、台地玄武岩とも呼ばれている。古生代カンブリア紀以後何回か発生している。

ナミビア北部に広がる洪水玄武岩
洪水玄武岩は、数百回以上繰り返し発生した噴火で形成された。
つまり流動性が良く、薄く拡がる玄武岩質溶岩が、何度も繰り返し噴火することで、現在見られるような、高大な台地や高原が形成された。この溶岩流を一枚ずつ分析すると、必ずしも同一成分で無いことが多い。
更にマントルを構成する鉱物群とは、かなり異なった成分の溶岩流が各地で観測されている。つまり洪水玄武岩は、マントルそのものが溶解したマグマが、継続的に噴火したものではないと考えられる。
最近の研究では、プレート運動で、海溝からマントルに沈み込んだ海洋地殻が、ホットプルームの上昇に巻き込まれ、地殻の下まで上昇し、そこで溶解して大量のマグマを形成したと説明されている。

エンディカ洪水玄武岩
洪水玄武岩は、比較的短期間に、非常に大量の溶岩が地表に噴出したため、当時の地球環境に重大な影響を与えた、と考えられている。玄武岩質溶岩の噴火では、火山灰の噴出量が少ない反面、大量の火山ガスを出すことが多く、この火山ガスが、環境に大きな影響を及ぼす。
近年の事例では、1783年のアイスランド・ラカギガル噴火(玄武岩質溶岩の大規模な噴火)で発生した火山ガスの影響で、北半球が数年間寒冷化した。
これは日本では、天明の大飢饉という形で現れた。洪水玄武岩の噴出に伴う、火山ガスの噴出量は、ラカギガル噴火の数十倍以上と見積もられており、当時の地球環境に、深刻な影響を与えた可能性がある。
実際、最大規模のシベリア台地玄武岩の噴火時期は、史上最大規模の大量絶滅が起こったP-T境界と一致しており、この時期の大量絶滅の原因の一つと考えられている。#目 次
白亜紀末(K-Pg境界)の大絶滅
白亜紀末の約6千500万年前(K-Pg境界)で、恐竜・翼竜・長頚竜・モササウルス類・アンモナイト類が、同時に絶滅した。この絶滅によって、海洋生物の47%の属と、16%の科が消滅した。さらに上記大型動物以外に、いくつかの微化石グループも消滅している。

ただし、アラモサウルスなどの、ごく一部の属は、これを生き残った可能性がある。
白亜紀末、中米のユカタン半島に、直径約10㎞の巨大隕石が、落下したことが確認されている。推定直径、約180㎞のクレーターを作った隕石の直径は、約10㎞と推定されている。隕石の落下速度は、秒速20~30㎞で衝突し、高さ300mの津波が発生し、浅海では千mの津波が襲ったと考えられる。
巨大な天体衝突が起きると、衝突地点周辺には、爆風や高温の蒸気雲が発生し、巨大津波も起きる。

衝突の爆風による破片は宇宙空間に達し、それが大気に再突入するとき大気を加熱する。その後、大気中に巻き上げられた大量の塵や、森林火災による煤が地球全体を覆って、太陽の光を遮りそれによって寒冷化の時代に入る。
このように、隕石の衝突の結果、地球が寒冷化して、恐竜などが絶滅したと考えられている。豪雨や津波や竜巻などの被害は、桁違いに大きく、またインド大陸で、現在のデカン高原を形成する洪水玄武岩の噴火があった。

こうした環境の激変によって、多くの生物は死に絶えた。
さらに海中のプランクトンのほとんどが滅した。理由は、ユカタン半島は、主に石灰岩と石膏でできていて、そこに巨大な隕石が衝突し、大爆発が起き、石膏の岩石のなかの成分が蒸発し、三酸化硫黄になり、激しく大量の酸性雨を地球全土に降らせた。これによって海水が酸性化し、プランクトンの石灰(カルシウム)の殻も溶かされ、ほとんど死滅したとされている。
この大量の酸性雨によって、恐竜などの大量絶滅が起きたと推測されている。つまり、この隕石衝突の場所が、もし石灰岩や石膏質の地質と異なる場所で衝突していたら、大量の酸性雨が降らず、爆発の影響も限定され、これほどの絶滅はおきなかつたかもしれない。#目 次
大型魚類の誕生
ダンクルオステウスは、約3億5千万年前のデボン紀の海に生息していた、大型の魚類である。板皮類(ばんぴるい)というグループに分類され、古代魚の中では最大の生物であった。
「魚の時代」と呼ばれるデボン紀の海で、ダンクルオステウスは最強の捕食者として、生態系の頂点に君臨していた。しかし、デボン紀後期に起こった大量絶滅で、地球上から姿を消した。

ダンクルオステウス想像図
体長は6~9mあり、最大級の個体では10mに達したと考えられている。大型の生物が、多く存在しなかったデボン紀の海では、ダンクルオステウスの体は非常に大きなものだった。大きな体を持つだけでなく、積極的に獲物を捕食する、獰猛な肉食魚でもあった。
デボン紀は魚類が大繁栄し、「魚の時代」とも呼ばれている。
それまでの海は、ウミサソリなどの、節足動物やオウムガイなどの軟体動物が支配していた。デボン紀に入ってから、魚類の巨大化が進み、捕食される立場から、捕食する立場に変わる魚が多く出現した。
ダンクルオステウスを始め、大型の魚類は、節足動物や軟体動物に代わって、デボン紀の海を支配するようになった。
ダンクルオステウスのように、体表の一部を、外骨格化させた魚類の仲間は、甲冑魚と呼ばれている。

甲冑魚の多くは、頭部を硬い骨板で覆われていた。これは強力な捕食者から急所となる頭部を守るためである。
それまで海で繁栄していた、節足動物や軟体動物の仲間には、頑丈な外殻や殻を持つものが多くいた。これに対抗し、魚類も硬い装甲板を持つようになった。
甲冑魚の仲間が増えたことで、それを捕食する生物の中には、装甲板を嚙み砕くための、強力な顎を持つものが現れる。
ダンクルオステウスは、大きな体に頑丈な装甲板を持ち、獲物の外殻を破壊する強力な顎まで持っていた。噛む力は5トン以上あったといわれている。
顎に歯は生えていないが、牙のように突き出した顎の骨自体が、その役割を果たし、強力な顎の力で、獲物となる生物の外殻を破壊していた。#目 次
大森林の誕生
4億5千万年前頃に誕生したシダ植物の段階では、葉・茎・根のしくみが構築され、土の上でも繁殖できるようになった。
この構造の獲得により、コケ時代の緑の絨毯が徐々に立体的な姿へと変わって行く。そしてついに3億5千万年前頃には、地球最初の森が誕生したと言われている。
これは現在のシダ類の仲間ながら、森を作り上げたシダの先祖たちは、現在のシダより遥かに巨大な木生シダだったのである。

裸子植物の大森林時代(石炭紀)
現在のシダ植物は基本的に草本であり、茎は這うかごく短く立ち上がる。そんな中、木生シダは、外見的に太い幹があり、ある程度の高さまで直立して育つものを指す。木性シダ、樹状シダともいう。
大きいものでは20mに達する例もある。これらは分類上は、シダ綱に属するもので、特にヘゴ科とタカワラビ科に多い。しかし、それ以外の群にも散発的に見られる。
また熱帯域を中心に見られる。茎はほとんどの場合に分枝せず、先端部に大きな葉を輪生状に出すので、外見的にはヤシ類やソテツ類のような姿になる。
しかしながら、この群では、茎に二次成長(肥大成長)をするための構造が存在しない。つまり、茎そのものは太くなってゆくことが出来ない。この類では、根元に近い太い幹は、その中心部に茎があるものの、大部分は茎の表面から、多数出て伸びた根が折り重なったものである。

石炭紀のリンボクの森林
なお、古生代には、広義のシダ植物に類するもので、大木になるものが出現し、それらは、茎に二次肥大成長を行う構造を発達させたものが含まれる。
これらも広義には、木生シダと言うが、狭義には、ヘゴ科に見られるような形のものだけを、この名で呼び、現生のものは全てこれに属する。
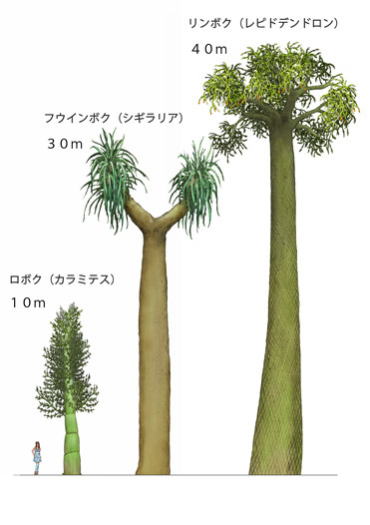
リンボクは、石炭紀の代表的なシダ植物の仲間で、沼沢地に群生し、1ヘクタール当り千~2千本も密生していたと考えられている。
リンボクは、伸びきるまで分岐しなかったので、この密集が可能となったといわれている。枝は二又に分かれ、細い枝には小葉が付いており、枝の先にはレピドストロブスと呼ばれる胞子嚢をつけていた。巨木を支えていたのは、スティグマリアと呼ばれる担根体(たんこんたい)である。リンボクは巨大になるが、木部は少なく、木部の外側に、厚い皮層を形成して幹を支えていた。幹の強度は小さかったと考えられ、現在の裸子植物のような樹木とはすこし違っていた。
現在、確認された最古の陸上植物は、チェコスロバキアのシルル紀から産出した、クークソニアだが、デボン紀(4億8千万年前)になると、多くの陸上植物が、パンゲァ大陸の各地から報告されている。しかしながら、これらのうちニューヨーク州で発見されたキャリキシロンという樹幹の化石を除いては、いずれも小型の草木性植物であった。
デボン紀に、高温多湿の熱帯性気候下にあった、欧米の一隅に発生したシダ植物は、葉・茎・根の仕組みを活用し他の植物より光合成に有利な「高さ」を獲得している。
そしてシダ植物に属する、鱗木・封印木・藍木などの、巨木からなる森林が生まれた。 この森林が、石炭紀に入ると、欧米を中心に、造山帯の低地や内海沿いの湿地帯に爆発的に大繁栄し、各地に大森林を形成して行ったのである。
その名残が、石炭として今でも多く残っている。石炭は、この時代の植物が、完全に腐敗分解する前に地中に埋もれ、そこで長い期間地熱や地圧を受けて変質したことにより生成された。いわば植物の化石でもある。

石炭紀の地層
石炭は、古くから燃料として使われてきた。特に産業革命以後20世紀初頭までは、最重要の燃料として、近代エネルギーの一つとして大きな役割を担った。
その石炭は、殆どが、この時代の森林を形成したシダ植物である。石炭紀に、広く発達したシダ植物の大森林は、その後、米国のアパラチア炭田、欧州の諸炭田、ソ連のポドモスクワ炭田やドネッツ炭田などの、大炭田を形成していった。そして、この時代が「石炭紀」と呼ばれる由縁である。
#目 次
巨大昆虫
この巨木の森林が形成した立体的な環境の中で、植物と共に上陸した昆虫たちも一気に繁殖を始める。大森林の3次元空間で、新たな機能を獲得し、ついに生物史上初めて空を飛ぶ昆虫が誕生したのもこの時代の、この森の中での出来事だった。
そして脊椎動物が上陸するまでの間、地上は巨大シダ植物と、昆虫の楽園として繁栄していった。 現在でも、昆虫類は、地球上で繁栄を極めており、500万種以上の種類があると言われている。

巨大トンボメガネウラの化石
ちなみに、現在のほ乳類は約4千種であり、昆虫はずば抜けた環境への適応力を持っている。さらに、この時代の昆虫は、巨大で80㎝のトンボや、6枚羽で最重量を誇60㎝のカゲロウや、2mを超えるムカデたちが活動していた。
昆虫巨大化の理由は、シダ植物の大繁殖による酸素濃度の上昇であるといわれている。しかし、シダ植物の水分運搬の仕組みはまだ不完全であり、しかも繁殖では水に依存する部分があったので、まだ水の豊富なところでしか繁殖できず、内陸に進出することは出来なかった。その後、巨大シダ植物は、環境の変化と共に、新たに登場してきた種子植物へ、交代して行くことになる。

アースロプレウラ想像図
アースロプレウラは、古生代石炭紀の森林に生息した、巨大な節足動物である。
節足動物とは、エビやカニなどの甲殻類、クモやダニなどのクモ形類、膨大な種数の昆虫類などの大きな群である。
節足動物の仲間では、ウミサソリの一種と並んで、史上最大級のものと考えられる。
ほぼ同じような形の、多数(少なくとも20節)の体節がつながった構造をしていた。全長は2m~3mに達し、幅は45㎝。ノバスコシア、 イリノイ、オハイオ、ペンシルベニアなど、各地から化石が見つかっている。完全な標本はなく、詳細には謎が多いが、多足類のムカデや、ヤスデに似ており、これらに近いものと考えられている。
ムカデは、小動物を捕食する肉食であり、ヤスデは雑食だが、アースロプレウラは植物食であったと考えられている。
たくさんの体節がつながった柔軟な体は、森林内を移動するのに適していた。
大きな動物で体重も重く、這った跡の化石が、各地で知られている。石炭紀に、アースロプレウラや、メガネウラのように、巨大な節足動物(昆虫も含む)が多かった原因は、シダ植物群の大繁殖により、当時の大気中の酸素濃度が約35%と高かったこと、脊椎動物が、陸上での進化を始めて間もない頃であり、これらを捕食する動物が少なかった事などの説などがある。
ペルム紀のはじめには、巨大な節足動物のどれもが絶滅した。
その原因は、捕食動物によるものではなく、気候の変化とともに、石炭紀と同じ森林環境が姿を消し、食料となる植物が無くなったことだとされる。 #目 次
爬虫類
爬虫類は、脊椎動物の分類群の一つで、分類上は爬虫綱という単位を構成する。現生ではワニ、トカゲ、ヘビ、カメ、ムカシトカゲが含まれる。爬虫類の「爬」の字は「地を這う」の意味を持つ。

古生代に地上で生活する生物群として、3億年前に両生類から進化した爬虫類は、急速に多様化した。そして爬虫類は、その前にいた両生類に代わり、世界を支配し始めた。
古生代半ばから中生代前半にかけて、哺乳類型爬虫類が、中生代には恐竜、翼竜などが、新生代からは鳥類や哺乳類が繁栄した。
現生種としては、通常はトカゲ類(ヘビ類を含む)、カメ類、ワニ類、ムカシトカゲを含み、鳥類と哺乳類は含めない。体表は表皮の変形した鱗でおおわれ、4本の脚と尾、乾燥に強い卵(有羊膜卵)などが特徴である。

また蛋白質の代謝によって発生するアンモニアは、両生類や哺乳類のような尿素ではなく、水に不溶である尿酸に代謝し、糞とともに総排出腔から排泄するものが多い。
これも乾燥に対する重要な適応の一つである。ヘビや、アシナシトカゲを始めとする一部のトカゲのように、脚が退化しているものや、カメ類のように鱗と骨格が一体化し、甲となったものもある。
繁殖形態は卵生で、革質か石灰質の殻におおわれた卵を、陸上に産む。
生まれた子供は、親と同じ姿をしており、変態はしない。ただし直接子供を産む卵胎生の種もいる。キノボリヤモリ、オガサワラヤモリ、ブラーミニメクラヘビなど、単為生殖種が一部に存在する。
多くは変温動物で、体温が外部温度に少なからず依存する。昼行性の爬虫類の多くは、日光浴等で体温を高めた上で活動を始める。

変温動物で体温を保つ必要がないため、哺乳類や鳥類に比べて、食事の間隔は長い。適度な水分さえあれば、何も食べずに1ヶ月ほど生きることもある。
現生の爬虫類の中には、ウミガメの一部の成体などのように、体温の変動幅がわずかな恒温動物的体温調節を行うものもいる。また哺乳類型爬虫類や、恐竜などの絶滅種の一部にも、恒温動物的体温調節を行っていたものが存在するのではないかと考えられている。#目 次
爬虫類の多様化
ペルム期末の大絶滅を生き残った、哺乳類型爬虫類から哺乳類が登場し、それとは別の爬虫類のグループから、ワニの仲間、カメの仲間、そして恐竜類、さらにワニ類に近い仲間から、空を飛ぶ翼竜類が登場した。
なお、哺乳類型爬虫類と哺乳類のことを単弓類、それからワニ類、恐竜類、翼竜類のことを双弓類と分類している。
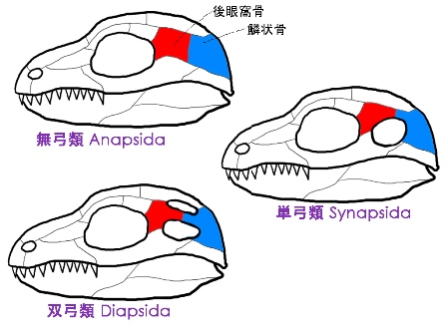
単弓類とは、目の後ろに穴が一つある爬虫類のこと。さらに細かく分類すると、
①盤竜類―初期の哺乳類型爬虫類。石炭紀後期に登場し、ペルム紀に大繁栄を経て絶滅。
②獣弓類―半直立姿勢ができ、頭骨が発達した哺乳類型爬虫類。ペルム紀に登場。となる。 さらに、この獣弓類の中で代表的なものが、ディキノドン類とキノドン類で、それ以外の獣弓類は、ペルム紀に殆ど絶滅している。
このうちキノドン類は、犬歯とほかの歯が分かれている異歯性を持ち、骨質の二次口蓋(鼻と口を隔てる骨)を持つことで、食べながら呼吸することが可能となったのが特徴である。そして多くは体毛があったと考えらている。
哺乳類の祖先は、このキノドン類から三畳紀に分化し、他のキノドン類は、三畳紀末から衰退し、ジュラ紀に絶滅した。
獣弓類のディキノドン
つまり獣弓類で、今も生き残っているグループは、哺乳類だけである。進化の系統を分岐学で見ていくと、哺乳類は、魚類から分化した両生類―分化した爬虫類―分化した哺乳類という系統になる。
双弓類とは、目の後ろに穴が二つある爬虫類のこと。さらに細かく分類すると、
①鱗竜類―有鱗目(トカゲ類とヘビ類)などが所属。このうちトカゲ類とヘビ類は、現在の爬虫類の種の95%以上を占めている。首長竜類も、この仲間に分類されるが、別に広弓類として同格に扱われることもある。
②主竜形類―主竜類(恐竜やワニ)、翼竜類などが所属。
③魚竜類。などに分類される。
三畳紀を代表するのが主竜類である。この主竜類は、さらに3つに分類される。
①恐竜類―恐竜のほか、恐竜から分化した鳥類も含む。
②クルロタルシ類―現世ワニ類の祖先とその仲間。
③翼竜類。と分類されている。 #目 次
恐 竜
恐竜は、脊椎動物の分類群の一つで、中生代三畳紀に現れ、中生代を通じて繁栄した。多様な形態と習性で適応放散し、陸上動物としては非常に大きくなったが、約6千600万年前の白亜紀と新生代との境で多くが絶滅した。
アラモサウルスなどの一部の属については、この後もしばらく生き延びていた可能性を主張する研究者もいる。一般的に恐竜の一群として印象されている翼竜、首長竜、魚竜などは恐竜には含まれない。ただし翼竜は、恐竜やワニと同じく、主竜類に属し、恐竜とは姉妹群にあたる。
首長竜や魚竜は、恐竜とは遠縁の水棲爬虫類である。
古典的分類では、爬虫綱 - 双弓亜綱 - 主竜形下綱に属し、分類階級は上目とされてきた。 なお、系統樹に基づく分岐学的観点から、単に「恐竜」と呼んだ場合、学術的には「鳥類」を含めることが多い。
恐竜は、多くの爬虫類とは異なり、胴体の直下に四肢を持つ。この特徴は、側方型の四肢に比べて体重を支えるのに都合がよく、大型化したグループが出現する素地となった。また歩行の際に、身体を捻る必要がないため、軽快な移動を可能にしている。
この特徴は、やや遅れて哺乳類も獲得しているが、異なる点としては、恐竜は二足歩行の種が多い点である。

これは、二足歩行が初期主竜類から受け継いだ、祖先的な形態だからである。
初期主竜類の中でも、ユーパルケリアなどは、身体の作りが軽快であり、常時二足歩行を可能としていた。竜脚類や鳥盤類の一部の様な四足歩行の恐竜は、体重の増加等の理由で、二次的に四足歩行に復帰したものである。このためか、四足歩行の恐竜でも、体重の大半は後足が支える形となっている。
恐竜の二足歩行は、ヒトとは異なり、後足を中心に長い尾によって、上半身と下半身のバランスをとっていた。

恐竜の巨大な尾は、バランスを取るための必然であり、ティラノサウルスや近縁の属に見られる、縮小した前肢は、巨大化した頭部と釣り合いを取るためだとされる。
一方、胴体の横から足が生えている側方型の爬虫類、例えばトカゲは、胴体をくねらせて歩行する、いわば爬行(はこう)を行っている。
この方法では、四肢を側方に突き出した姿勢で、身体を持ち上げているため、エネルギー効率が悪い。
また肺を圧迫するために呼吸が阻害され、長時間の走行を困難にしている。
カメは四肢が側方に生えていながらも、胴体が甲羅に覆われて可動性を持たないため、爬行を行えない。そのため四肢で胴体を持ち上げて歩行を行っているが、歩行能力は劣って いて、歩行速度の遅い動物の代表格扱いされている。

三畳紀のワニ
またワニは、トカゲに比べて、胴体の可動性がやや乏しいために、胴体をくねらさない。また、短距離ではあるとはいえ、ギャロップで走行することが可能なものも存在する。ただし、ワニは恐竜に近縁なグループであり、祖先は胴体直下に直立した四肢を持っていた。そのため、かれらの四肢を突き出した姿勢は、半水生の生態に適応した二次的なものである。 またカメもワニなどに近縁な主竜形類であるため、祖先は胴体直下に四肢を持っていたが、甲羅の発達にともない、二次的に四肢が側方へと突き出す形となったと推測されている。
恐竜の二足歩行形態は、現在では子孫の鳥類へと受け継がれている。しかし、祖先と異なる点としては、尾が短縮したことで、重心が前方へと移ったため、大腿骨がほぼ身体に対して水平に保持されている。そのため、歩行は膝関節を中心としたものとなっている。
#目 次
巨大な恐竜の誕生
恐竜は、哺乳類と同時期に爬虫類が、地球の陸地が乾燥するにつれて、乾燥した環境に順応して体を進化変化させていき、ついに恐竜は気嚢を発達させ巨大化し、2億年ほど前に恐竜へと進化を遂げた。
この時期に、恐竜や哺乳類が誕生した背景の一つは、大気の酸素濃度が低い環境になったためと考えられている。哺乳類は横隔膜を、恐竜は気嚢というものを発達させて、低い酸素濃度でも活動できるように進化した。
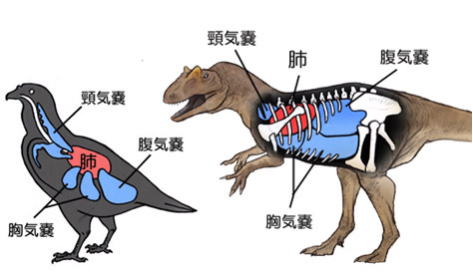
恐竜や哺乳類が誕生した最初は、哺乳類の方が優勢だった。
これが逆転した理由は、恐竜の気嚢システムは、骨格を軽くする事ができ、この理由で巨大化には哺乳類より有利だった。
気嚢システムは、恐竜の生き残りといわれる鳥類が、今も受け継いでいる。骨格を軽くできる特徴は、当然飛ぶのに有利である。また、鳥の中には、ヒマラヤ山脈を飛び越えるものがあるが、これは薄い空気でも呼吸可能な気嚢の利点による。
もう一つ、恐竜が巨大化で有利だったのは、草食恐竜が食べる植物の栄養が、あまり豊富でなかった。 ―と言われている。
このため、草食恐竜は多くの植物が必要で、そのために巨大化し、それを食べる肉食恐竜も、それに合わせて巨大化していったと考えられている。
その進化した恐竜が地球全体に広がり、やがて陸・海・空すべてを支配するようになった。そして恐竜が6千500万年前に絶滅するまで、1億4千万年もの長い間、恐竜の時代が続いた。
最初の三畳紀で、恐竜、翼竜、魚竜、首長竜、ワニ、カメなどが誕生し、最古の哺乳類も誕生した。三畳紀の後期には、急速に進化を遂げ、恐竜繁栄の時代が到来した。そしてその他の大型動物が、激しく生存競争を繰り返した。
#目 次
両生類の誕生
両生類は、古生代の石炭紀頃以降、多くの化石種が知られている。
しかしながら、現在でも生息しているものは、長い尾を持ち、短い四肢のある有尾目のサンショウウオなどと、尾がなく体幹が短くまとまって四肢の発達した無尾目のカエル類、それに四肢を失い、細長い体の無足目のアシナシイモリ類の3群のみである。

サンショウウオ
両生類は約3億6千年前、陸上での生活も始めたと考えられている。これが脊椎動物の中では初めて陸上生活が可能となったと考えられている。
ただ陸上生活が可能とは言っても、その身体の構造、生活史、生理、生殖などで、陸上生活への適応を示しながらも不充分であり、水辺への依存度が強い。特に幼生は、一般に水中生活をするなど、基本的に水中環境が欠かせない。
現生の種は、ほぼ全てが淡水域を生活の場としている。
原始的な形では、水中で産卵し、幼生は四肢を持たない形で生まれ、鰓呼吸で水中生活を行う。その後変態を経て、肺呼吸で陸上生活の出来る成体になる。

ただし、多くの例外があり、その生活は多様である。ただ、基本的に乾燥に弱いため、水辺などの湿った環境が生息域の中心であり、陸上で活動可能な体を持ちながら、生活や繁殖を水に依存した生涯を送ることからこの名がある。
「両生」類の名は、水中生活と陸上生活の、両方が可能という意味ではなく、両方の環境が必要な動物である、という意味である―これが近年の両生類の減少に繋がっているとの指摘もある。本来、両棲類、両棲綱であったが、現在は多くの場合「両生類」「両生綱」と書かれる。 #目 次
大型草食性恐竜
アパトサウルスは、約1億5千万年前(中生代ジュラ紀後期)の北アメリカ大陸に棲息していた、竜盤目― 竜脚形亜目― 竜脚下目 ― ディプロドクス科の一種である。
形態と生態は、全長は約21~26m、体重は推定方法によって幅があるが、約24~32トンという見積もりがある。
群れを成して移動、森林の木の葉を常食していたと考えられる。1960年代までは、あまりに体重が重く、脊椎骨に多くの空洞があって、重心が低位置にありまた首が長いこと、鼻孔が頭の上部に開口していることなどが、水中生活と推測されていた。
その後の研究で、陸棲であったことが判明した。
アパトサウルス
竜脚類のような大型の恐竜では、死後、堆積物に埋没されるまでに時間がかかるため、骨がばらばらの状態で化石化することが多く、特に頭蓋骨は、早い段階で失われてしまう。
アパトサウルスの場合も、完全に近い骨格標本が得られていたにもかかわらず、頭骨のみ、長い間発見されなかった。
1990年代、より完全な頭蓋骨の発見で、アパトサウルスが、ディプロドクスと近縁であることが判明。ディプロドクスを参考にした頭蓋骨で復元されるようになった。
細長いディプロドクスと異なり、カマラサウルスのような、ややずんぐりした体形だが、ディプロドクスと共通の鞭のような尾を持っていた。化石はアメリカのジュラ紀下部のモリソン累層から発見されている。

アパトサウルスの化石標本
約1億5千万年前のこの地域は、単独の大陸として、北半球に位置した北アメリカ大陸の西部地域である。白亜紀に、この大陸を東西に分断することになる。
ディプロドクスは、約1億5千万~約1億4千700百万年前(中生代ジュラ紀後期)の、北アメリカ大陸に生息していた、大型草食性恐竜の一種。
属名は「地震トカゲ」の意で、「歩くと地震が起きるほどの巨体」ということから命名された。推定全長約20~35mで、アパトサウルスなど他の竜脚類と比べ華奢で、体重は比較的軽く、10~40トン程度と見られる。

最大の特徴は、細めの体と長大な尾である。この尾は、強靭な筋肉によって、鞭のように振るなど、主に自衛のために使われたと思われ、全力で打ち振るわれた尾の先端部は、鞭のように音速を超えたという説もある。
四肢はその巨体に比してやや短く、腰側がやや低いため、胴体は後傾する。巨大な植物食動物で、足跡の化石などから、他の竜脚類と同様に、群れを成して生活していたと考えられている。
頭蓋骨は高さが比較的無く、前後に長く、口の前半に鉛筆のようなまっすぐな歯が生えていた。他の竜脚類では、スプーン状の歯を持つものが多く、この形はディプロドクス科に固有の特徴である。
より進化したティタノサウルス科も、この形に近い頭蓋骨と歯を持つため、一時期ディプロドクス科と近縁と考えられたが、初期の種の特徴から、むしろブラキオサウルスに近いことが分かり、収斂進化の結果と見なされる。
この独特の歯で、樹木の葉を櫛で梳(す)き取るように食べていたと、推測される。他の竜脚類と同様、噛まずに呑み込まれた葉は、筋肉質の胃袋の中で、胃石とともにすり潰されて消化されたようで、化石の腹部から、磨耗して丸くなった石が多数発見されている。
1979年、発掘された骨格は、現在この一つのみで、発見当初は、全長50~ 60mとも言われていたが、現在は前述の体格と判明した。当初は、史上最大の恐竜とされていたが、1993年にさらに巨大とされるアルゼンチノサウルスが発見され、その座を譲った。 それでもスーパーサウルスや、マメンチサウルスと共に、ジュラ紀最大級の恐竜である。#目 次
最大級の肉食恐竜
ティラノサウルスは、約6千850万~約6千550万年前(中生代白亜紀末期)の、北アメリカ大陸に生息していた肉食恐竜。大型獣脚類の1属である。
属名 テラノサウルスは、古代ギリシア語のテュラス ―「暴君」 + サウロス―「とかげ」の合成語で、「暴君の爬虫類」といった意になる。

ティラノサウルス
現在、史上最大級の肉食恐竜の一つに数えられ、地上に存在した最大級の肉食獣でもある。恐竜時代の最末期を、生物種としては約300万年間も、生態系の頂点に君臨したが、白亜紀末の大量絶滅で最期を迎えている。
非常に名高い恐竜で、最強の恐竜として描かれることが多い。また恐竜時代終焉の象徴として、滅びの代名詞にも度々引用される
骨格標本から成体の体長は、約11~13m、頭骨長は約1・5mで、体重は概ね5~6トンと推測されている。 ティラノサウルスの化石は多くはなく、2001年の時点で、20体程で、完全なものは3体のみである。

ティラノサウルスの上下の顎には、鋭い歯が多数並んでいるが、他の肉食恐竜と比べると大きく分厚くて、最大で18㎝以上にも達する。また、餌食となった恐竜の骨の多くが、噛み砕かれていたことから、驚異的な咬合力を持っていたと考えられる。
その咬合力は、少なくとも3トン、最大8トンに達したと推定されている。これらの事実から、小・中型獣脚類が爪を武器として用いていたが、ティラノサウルスは、強大な顎と歯のみを武器として使用していたらしい。
ティラノサウルスは、各部位によって僅かながら歯の分化が進んでいたとされる。特に門歯は、断面が特徴的なD字型をしており、ティラノサウルス類を見分ける上での指標になっている。
頭蓋は同じ大きさの他の獣脚類に比べて、明らかに幅広であり、特に後眼窩部の張り出しが著く、吻部も丸みを帯びた広い形になっている。脊椎骨数は、頚椎:10、胴椎:13、仙椎:5、尾椎:35~44。頚椎、胴椎といくつかの仙椎は側腹腔)を生じ、椎体には、含気化が進んでいる。

体の大きさに比べて前肢は異常に小さく、指が2本あるのみで、用途は未だにはっきりとしていない。逆に頭部は非常に大きく、それを前肢の代わりに、上手く活かしていたのではと考えられている。
また、進化の過程で、体の前方が重くなったため、前肢を短く軽くすることでバランスを取ったとする見解もある。
ティラノサウルスとその類縁種は、足の速いオルニトミモサウルス類(ダチョウに似た恐竜群)と、共通の特徴であるアークトメタターサルを有していた。
アークトメタターサルとは、第三中足骨が、第二、第四中足骨によって挟み込まれ、上端が押しつぶされる形態のことを指す。#目 次
哺乳類の時代
K―T境界で起きた生物の大規模絶滅事件で、中生代に地上・海中・空中に繁栄していた恐竜などの、大型爬虫類は、ワニ類を除いてほとんど絶滅した。
その結果、生態的空間を埋めるように、生き残った哺乳類が、適応放散して陸上から海中にも進出し、様々な環境に適応して種類を増加させた。このため、この時代を哺乳類の時代とも呼ぶ。
一方で、多様性の数からは、昆虫類のほうが多様性は高い。
新生代は、哺乳類と鳥類の適応放散が起こった時代。
鳥類は、すでに白亜紀で空中でも陸上でも、翼竜や恐竜と伍して生活しおり、哺乳類より有利であり、古第三紀最初(暁新世)の、最大の捕食者は鳥類のディアトリマであった。
ところが、やがて進化した哺乳類との生存競争に敗れ、このような大型の地上棲鳥類は衰退した。
一方で、白亜紀末の大絶滅を生き残った小型の鳥が、より現代的な姿へと進化し、特にスズメ類、オウム類、キツツキ類などが充実してきた。空を飛ぶ動物といえば、これとは別に、最初のコウモリ類や、現世の蛾や蝶の祖先も現れている。

アデロバシレウス
古第三紀初期の哺乳類は、ほとんどが草食や昆虫食で、大きさもネズミほどのものが多く、最大のものでもネコ程度であった。
しかしやがて多くの爬虫類が絶滅した地上に適応し、やがて体も大きくなってゆく。
哺乳類は、暁新世から始新世にかけて、一次適応放散の後、漸新世で2度目の適応放散が起こった。現在見られる哺乳類の多様性は、漸新世から始まった。すなわち現代型のクジラ、げっ歯類のリス・ネズミ、長鼻類のゾウ、霊長類の真猿類、奇蹄類のウマやサイ、偶蹄類のイノシシやラクダ、食肉類のサーベルタイガーやクマなどが、漸新世に現れた。
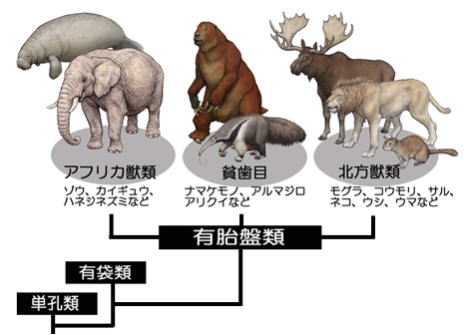
なお 新生代初め、オーストラリア大陸は、南極大陸とつながって、他の大陸とは海を隔てていたため、これらの哺乳類(真獣類)とは、系統が異なる単孔類や有袋類が適応放散していた。
オーストラリア大陸の生物の特殊性は、人類がオーストラリアに渡るまで継続した。同じように、他の大陸と離れていた南アメリカには、一部の真獣類と有袋類が繁栄した。#目 次
始祖象の誕生
メリテリウムは、長鼻類で、ゾウの祖先に近い仲間。 長鼻類は、アフリカ大陸に起源を持つアフリカ獣類に属し、テティス海の岸辺(アジア大陸南端地域とアフリカ大陸北地域)で進化的飛躍を遂げたテティス獣類に含まれ、ゾウ(象)とその近縁群を含む。
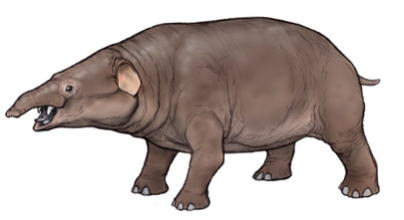
メリテリウム― 始祖象
メリテリウムは、ゾウの仲間である長鼻類の一グループ(属)で、ゾウの先祖の一つとされ、始祖象ともいう。
北アフリカのエジプト、アルジェリア、マリ、リビアの各地で、始新世後期から漸新世初期 ―約4千万~3千万年前の地層から、 化石が発見されている。20世紀はじめ、最初に発見されたカイロ南西、ファイユーム盆地の古い湖のメリ湖の名前から、メリ(Moeri)の獣(therium)という名前がつけられた。

メリテリウムの化石標本
いまのゾウとは異り、肩の高さは70㎝ほどで、大形のブタか、小形のバクぐらいの大きさで、頭はやや長く、胴長の哺乳動物であった。鼻は短 メリテリウムの骨格化石 かったが、切歯は大きく牙状であり、全体の体の形は、現生のコビトカバによく似ていて、水陸両用の生活をしていたと考えられている。
ゾウの先祖を含む古い哺乳類のグループの一つとして扱われている。体の構造から、系統的には、絶滅哺乳類のデスモスチルスのような束柱類、ジュゴンなどの海牛類、ハイラックスなどの岩狸類に系統的に近いともされていて、それらに共通な先祖のなかにゾウの起源が求められている。メリテリウムのような ゾウの先祖の仲間には、北アフリカから出土する、化石として知られているものが多い。

デイノテリウム
一番古いものは、およそ6千万年前の暁新世後期の地層から見つかった、フォスファテリウムがあり、これは長鼻類の最古の先祖であるとともに、有蹄類全体の先祖であるとされている。
このほか、ヌミドテリウム、バリテリウム、デイノテリウムなどがある。それらは断片的な化石しかなく、長鼻類のなかでの位置づけや、相互関係については明らかでない。
なお、パキスタンやインドで化石として知られているアントラコブネというグループにも、そのような長鼻類の先祖の一つがあるとされている。#目 次
ウマ科動物の誕生
メソヒップスは、奇蹄類の中でも初期に登場したウマ類。
その起源は、約5千500万年前に遡る。メソヒップスは、その中でも最初のウマ類で、木の葉を食べていた。

メソヒップス
ウマ科を含む奇蹄目は、K―T境界の後、1千万年までの暁新世後期に誕生した。
奇蹄目は元々、熱帯林での生活に順応していたが、バク科とサイ科が森に適応したのに対し、ウマは草原などステップ地帯での生活に適応した。その過程で、次第に背が高くなり、足指では中指の発達と平行して、他の指の退化が進むなど、一定方向への系統的な変化がみられる。
最も古いと考えられるウマ科動物は、ヒラコテリウムである。
ヒラコテリウムの化石は、18世紀にヨーロッパで発見され、「ハイラックス様の獣」を意味する、ヒラコテリウムと名づけられ、正式学名も「ヒラコテリウム」となっている。
ヒラコテリウムは、約5千200万年前に、北アメリカ大陸で生活していたとされている。体はキツネと同じくらいのサイズ、―体高25 ~45㎝で、比較的短く弾力性のある頭頸部とアーチ状の背骨を持っていた。
歯は各側に切歯を3個、犬歯を1個、小臼歯を4個、大臼歯を3個備えており、合計で44個の歯を持っていた。
大臼歯は、葉を削りやすい形であり、ヒラコテリウムは葉食性 ―柔らかい木の葉や果物などを食べていた。

またヒラコテリウムは、小さい脳を持っており、小さい前頭葉もあった。すでに走ることの進化は始まって、手足は現在の馬のように、体に比例して長かった。しかし、下肢骨のいくつかは不安定で、柔軟性に欠けていた。
脚はそれぞれ5本ずつ指があったが、進化の過程で、前肢は第1指が退化し4本、後肢は第1指と第5指が退化し3本になっている。
爪先は犬のような鉤爪ではなく、小さなひづめがついていた。
約200万年の間に、ヒラコテリウムは進化し繁栄した。
最も重要な進化が、より葉食性に特化した歯の獲得である。始新世の間、ヒラコテリウムは、ウマ科の様々な種に分岐した。
これらの完全な化石は、北米(ワイオミング州ウィンド川など)で数多く発見された。現代のウマの先祖とは考えられていない、プロパラオテリウムなどの化石がヨーロッパでも見つかっている。プロパラオテリウムは、パラオテリウムへと進化するが、その後絶滅した。
#目 次
第三紀の動物
新第三紀の前半の中新世に、現代の哺乳類のほぼすべてのグループが出現した。
サイやウマなどの奇蹄類、ラクダなどの偶蹄類といった哺乳類も出現していた。 奇蹄類は、サイに近い仲間やウマの仲間が生息していたが、衰退してゆく。
また種の数や個体数も、現在よりも多かったとみなされている。

海中では、クジラ類からイルカ類が生まれ、樹上生活の真猿類の中から、類人猿が現れた。偶蹄類の適応放散が進み、イノシシ、ラクダ、シカ、ウシ、キリンが、オーストラリアと南アメリカを除く世界中に広がった。長鼻類のマストドンも、現在のゾウの分布よりはるかに広い範囲に生息した。
食肉類は、イヌ、ネコ、イタチ、クマがそろった他、アシカ、アザラシ、セイウチなどが生まれた。この真獣類の繁栄は、新第三紀後半の鮮新世にも続き、ほぼ現在見られる動物と同じタイプの生物がそろった。約350万年前に、パナマ地峡ができて、それまで他の大陸から離れていた南アメリカ大陸と、北アメリカ大陸がつながった。
それまで南アメリカで繁栄していた有袋類は、オポッサムを例外として、北アメリカからやってきた真獣類との生存競争に負けて姿を消していった。
植物界では、約700万年前に、新しい光合成システムを持つ植物が現れた。光合成はシアノバクテリア以来、カルビン回路と呼ばれる合成方法が唯一のものであったが、低濃度の二酸化炭素を、効率よく利用できるC4型光合成を有するトウモロコシやサトウキビが生まれた。#目 次
種の形成
種とは、生物分類上の基本単位である。
現在命名済みの「種」だけで200万種あり、実際は、その数倍から十数倍以上の種の存在が推定される。
新しい種が形成される現象、メカニズムを種分化という。
生物は、無数の個体からなり、それらが非常に多様な形質を持つと同時に、一定の類型に分けられ、それらに名前を付けていた。たとえば虫、魚、鳥、草、苔などである。
更にそれらの大まかの分類の中にも、多様な形質を観察することで、より細かい不連続な集団に分けられた。つまり、形質のかなり細部までが共通する集団が見分けられ、それらの集団の間には、不連続性が見られる。
たとえばミカンの木につく青虫を育てれば、そこから出てくるチョウは、黄色のまだらのものか、真っ黒の羽根のものかである。
前者はアゲハチョウで、後者はクロアゲハだが、それらは色だけでなく、羽根の形や幼虫の姿でも少し異なっている。また、このような形質は、世代を越えて維持される。
そのような集団を種という。
博物学や生物学の知識の蓄積に伴って、すべての生物が、このような集団に区分できることが明らかになった。それぞれの種に、体系的に名を付け、分類体系を築こうとしたのがリンネである。
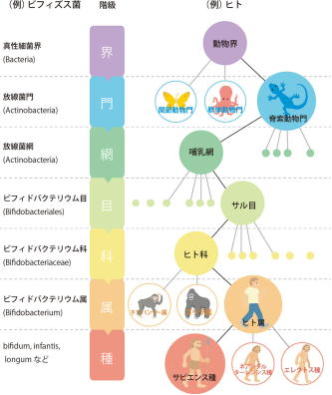
その百年後には、ダーウィンが進化と種分化の理論を提唱し、リンネの「形態に基づく分類体系」が、何故そのようになっているか、理論的説明を与えた。しかし、リンネの時代には、生物はまだ真核生物しか知られていなかった。
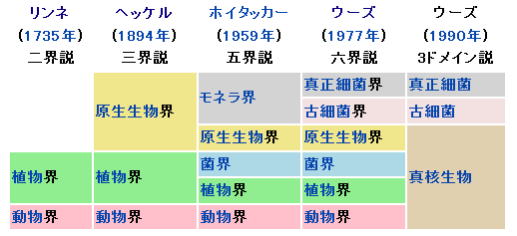
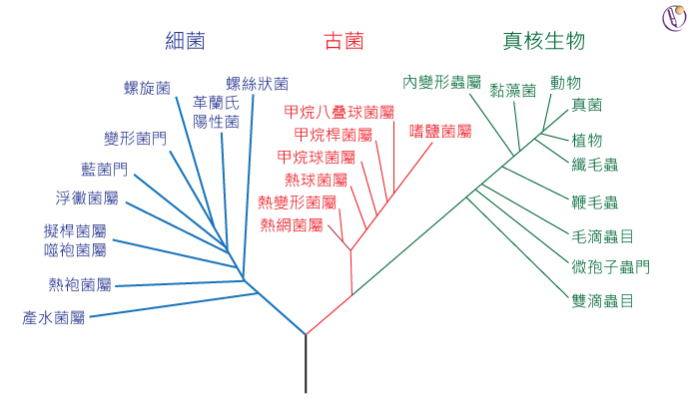
現在それ以外にもモネラ、真正細菌、いわゆる狭義の細菌、アーキア、古細菌、そして生物かどうかの異論もある、ウイルスやウイロイドといった存在があることが知られている。そして、いわゆる真核生物とはモネラ(生命初原物質)とアーキア(古細菌)、見方によってはウィルスが複数共生した複合生命体であることが定説になっている。

このため、リンネの考えた種の概念は、真核生物では比較的よく適合するが、それ以外のモネラ(真正細菌)、アーキア(古細菌)、ウイルスやウイロイドといったものには適合性が良くない。
モネラ、アーキアはリンネの唱えた2名法による種名が付いているが、その概念と範囲は真核生物における物とは全く異なることに留意すべきである。ウイルスやウイロイドではそもそも2名法による種名は付けられていない。
個体間で生殖が可能かどうかは、種の判断で重視される。
これは、種の特徴が、世代を越えて維持されるものであること、古くは同種であれば子供を残せるはず、との素朴な判断があったためである。
しかし、有性生殖は、それぞれの個体の属する系統の間で、互いの遺伝子を交換し合う行為であり、互いに交配可能であれば、いつかは実際にその遺伝子が、交換される可能性がある。そのような関係で結びついた個体の集団は、同じ遺伝子プールを形成する。
同一範囲の遺伝子集団を所有する限りは、形態的にもその同一性が保証されるはずと考えることができる。
しかし種における重要な概念の、「有性生殖(による遺伝子交換)」そのものが、真核生物に特有の概念である。
例えば、真正細菌では、有性生殖にあたる接合だけではなく、プラスミドの交換などを通して、相当に遠縁でも遺伝情報の交換ができる。
接合が知られていないものも極めて多く、相当遠縁同士でも、接合が起こることがある。また、外形は極めて変化に乏しいが、遺伝的には極めて多様なことが知られている。つまり、リンネの定義では、種を非常に細かく分けることも、非常におおざっぱに分けることもできてしまう。
DNA螺旋
現在の細菌の種の定義は、真核生物の分類と比較すると、非常に大きい集団を指しているものと思われる。例えば細菌の種分類の基準として、用いられることの多いDNA-DNA分子交雑法で、再結合率が70%以上であることや、核酸塩基配列の相同性が90%程度などを用いた場合、動植物では、目レベルの分類群が、全て同一の種に属することになる。 種の定義・概念は、現在、22以上あり、研究が進むほどに増加している。
#目 次
進化論
進化論(evolution theory)とは、生物が進化したものだとする理論、あるいは進化に関する研究や議論のことである。つまり、生物の種は不変ではなく、長期間かけて次第に変化してきた、という学説である。さまざまな生物は、全てその過程のなかで生まれてきた、とする説明や理論群である。

進化が起こっているということを認める判断と、進化のメカニズムを説明する理論という2つの意味がある。なお、生物学における「進化」は、純粋に「変化」を意味するものであって、「進歩」を意味せず、価値判断について中立的である。
進化は実証の難しい現象で、生物学のあらゆる分野から、進化を裏付ける証拠が提出されている。
初期の進化論は、ダーウィン仮説のように、画期的ではあったが、事実かどうか検証するのに、必要な証拠が十分に無い主張されていた。が、その後の議論の中で、進化論は揉まれて改良されつつある。現代的な進化論は、単一の理論ではない。
それは適応、種分化、遺伝的浮動など進化の様々な現象を説明し予測する、多くの理論の総称である。 現代の進化理論では、「生物の遺伝的形質が世代を経る中で変化していく現象」だと考えられている。
英語の evolution という語は、元来ラテン語起源で、内側に巻き込んでいたものを、外側に展開することを意味している。
従って、現在でも「展開」の意味で使われる。
最初にこの概念が生物学に援用されたのは、発生学の前成説においてであり、個体発生に際して、「あらかじめ用意された個体の構造が、展開生成する」プロセスを指していた。今日の日本語で、「進化」と翻訳されるような、系統発生のプロセスを指す語としての
evolution は、個体発生のこの概念を、系統発生に対して援用したものである。 みずからの進化論において、定められた方向への「進歩」を意図していなかったチャールズ・ダーウィンは、当初かれ自身は、この語を積極的に採用していない。
19世紀頃は、進化は進歩と同義であった。
その頃のヨーロッパでは、フランス革命や啓蒙思想などの普及によって、人間社会が発展のさなかであり、多くの人が、それが生物の進化と同じものであると主張していた。 それは神による創造の原点こそが最高の状態で、歴史のプロセスは、そこからの堕落による神からの離反であると、とらえるキリスト教的生命史観のもたらす不安からの、救済思想でもあった。20世紀には、社会的文化的変化が進歩と厳密に同義であるという考え方は、社会科学者から受け入れられないものとなっている。

また現在では一般的に、ダーウィンの進化の説明の解釈についても、生物の変化は進歩とは異なるものとして捉えられている。
19世紀以降は、進化と言った時は、社会や文化のそれでなく、生物の進化を指す。この生物の進化とは、ある生物の集団が、ある世代から次の世代に代わるときのアリル頻度の変化を意味する。
同一遺伝子座に属し、互いに区別される遺伝的変異体をアレルと呼ぶ。一つの集団内に存在する割合を、アレル頻度と呼ぶ。
アリル頻度は人種により大きく異なる。
それは、チャールズ・ダーウィンの自然選択のアイデアに基づく、種の進化論そのものでもある。「進化」は、単に事実を記述する語に過ぎないのであって、それ自体が価値判断を含むわけでなく、その意味で「進歩」とは異なるのである。にもかかわらず、進化と進歩の混同、事実と規範の混同はしばしば見られる。#目 次
チャールズ・ダーウィンの進化論
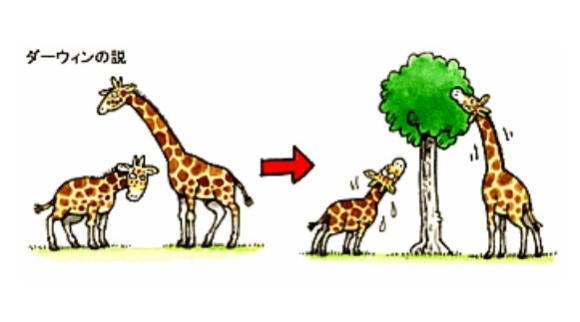
チャールズ・ダーウィンは、1831年から1836年にかけてビーグル号で地球一周する航海をおこなった。航海中に各地の動物相や植物相の違いから、種の不変性に疑問を感じ、ライエルの『地質学原理』を読んだ。そして地層と同様、動植物にも変化があり、大陸の変化によって、新しい生息地が出来、動物がその変化に適応したのではないかと思った。

1838年にマルサスの『人口論』を読み、自然選択説を思いついた。 ―と自伝には書かれている。
ハトの品種改良についての研究で、さらに考えがまとまっていった。1858年にアルフレッド・ウォレスが、ダーウィンに送った手紙に、自然選択説と同様の理論が書かれていたことに驚き、自然選択による進化理論を共同で発表したダーウィンは、さらに『自然選択』の要約をまとめ、1859年に『種の起源』として出版した。

『種の起源』のなかでは、現在の「進化」を指す用語として、あらかじめ内在的に用意された、構造の展開出現を意味する"evolution"ではなく、「変更を伴う由来」(Descent with modification)という語を使っている。
また自然選択(natural selection)、存在し続けるための努力(struggle for existence、現在では通常生存競争と訳される)、そして後の版では、ウォレスの提言を受け入れ、自然選択をわかりやすく説明する語として、ハーバート・スペンサーの適者生存を使用した。
これらの要因によって、環境に適応した形質を獲得した種が分岐し、多様な種が生じると説明した。
ダーウィンの説の重要な部分は、自然淘汰(自然選択)説と呼ばれるものである。
それは以下のような形で説明される。
生物がもつ性質は、同種であっても個体間に違いがあり、そのうちの一部は親から子に伝えられたものである。環境収容力は、常に生物の繁殖力よりも小さい。
そのため、生まれた子のすべてが、生存・繁殖することはなく、性質の違いに応じて次世代に子を残す期待値に差が生じる。
つまり有利な形質を持ったものが、より多くの子を残す。それが保存され蓄積されることによって、進化が起こる。
生物の地理的分布や、異性間に起きる選択である性選択についても説明した。
当時はまだ、 DNA や遺伝の仕組みについては知られていなかったので、変異の原因や遺伝については、うまく説明できなかった。ダーウィンの遺伝理論は、パンジェネシス―パンゲン説と呼ばれ、獲得形質の遺伝や、当時主流であった融合遺伝を認めていた。また発生と進化を明確に区別していなかった。
変異は、ランダムな物であると考えた。ここで言うランダムとは、「規則性が全く無い」と言う意味ではない。
ダーウィンは変異について、確実なことを述べられるような知識を何も持っていなかった。変異がランダムであるとは、変異それ自体には、進化の方向性を決める力が内在しないと言う意味である。進化を進歩とは違うものだと認識し、特定の方向性がない、偶然の変異による機械論的なものだとした。
「自然は跳躍しない」という言葉で、進化は漸進的であると主張した。 これは「進化は一定速度で進む」事を意味しない。
文字通り跳躍的な進化を否定するのみである。進化は小さな遺伝的変異の蓄積によって起きる。その結果として、体節数の変化のような、大きな形態的変化が起きる可能性はあるが、目や脳などが、一世代でできることはない。
一つあるいは少数の祖先型生物から、全生物が誕生した。そして一つの種が、二つに分かれる過程を、種分化と呼んだが、種分化のメカニズムに関しては深く追求しなかった。
ダーウィンは、進化の概念を、多くの観察例や実験による傍証などの実証的成果によって、進化論を、仮説の段階から理論にまで高めたのである。
ウォレスは、性選択説を認めず、非適応的と思われる形質(例えばクジャクの羽)も、自然選択で説明しようと試みたが、これは現在の優良遺伝子説に近い説明であった。
またウォレスは人間の高い知性や精神的能力は、神のような超自然的存在の干渉によるものだと考えた。 #目 次
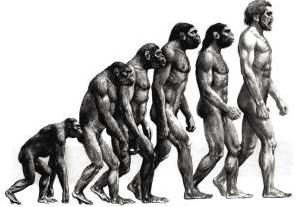
ホミニンの誕生と拡散
第四紀は、人類の時代とされる。
他の地質時代が、生物相の大幅な変化 ―特に大量絶滅を境界として定められたのに対し、第四紀は、人類の時代という意味で決められた。したがって、古人類学の進展に伴い、次々に古い原人が発見されるとともに、第四紀の始まる年代も変化していった。現在ではヒト属の出現を基準とし、地質層序や気候変動を併用して決定している。
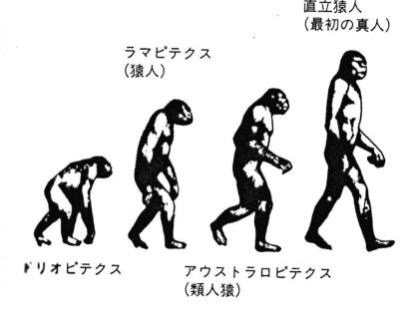
人類は樹上生活していた霊長類のうち、約700万年前にチンパンジーと枝分かれしたことが分っている。
そこからさらに派生した人類の仲間達は、23種、その中の一系統が、人類の進化の足取りである。23種の類人猿のほとんどは、新たな種を生み出すことはできずに滅んだ。
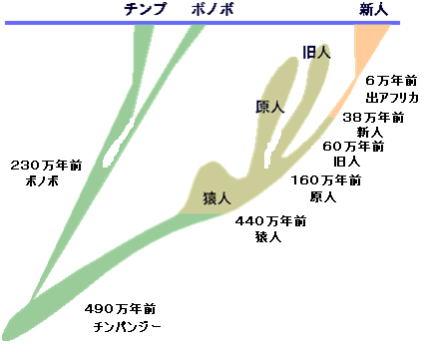
あるいは、ホモサピエンスの祖先によって滅ぼされた可能性もある。ともかく、アフリカに住んでいた類人猿から、現在のホモサピエンスは派生した。
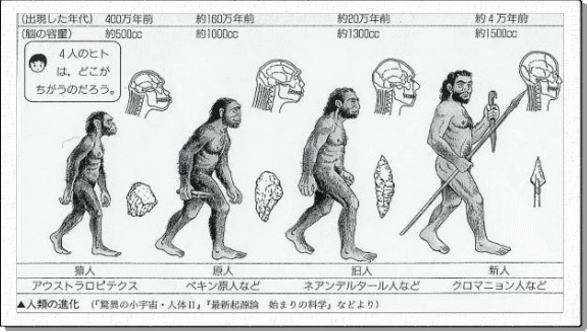
約440万年前のエジプトの地層から、類人猿と分かれて直立二足歩行した、ラミダス猿人の化石が、日本の調査隊によって1992~93年に発掘され、その後、ラミダス猿人の亜種は、約580万年前まで遡ることが判明した。
ラミダス猿人の次に、アウストラロピテクス(アファール猿人)が登場する。アウストラロピテクスの化石は、エチオピアや南アフリカの約250万年前~350万年前の地層から見つかっているが、骨格化石や足跡の化石から、確実に二足歩行していたことが確認された。歩行から開放されたアウストラロピテクスの手は、物をつかんだりする以外に、石を加工して石器を作ることができるようになった。
アファール猿人から、2種の猿人が派生した。
硬い植物を食べるために、頑丈な顎を発達させた猿人と、動物食で石器を活用し、脳を発達させた猿人である。
前者は約百万年前にすべて絶滅してしまい、後者の系統のホモ・ハビリスが現在の人類に続いている。
ホモ・ハビリス 脳容積は600㎜リットルで、チンパンジーの300~400㎜リットルよりはるかに大きい。
 
ホモ・ハビリス ホモ・サピエンス
次のホモ・エレクトスは、脳容積をホモ・ハビリス850㎜リットルに増やし、生存場所もインドネシア(ジャワ原人約20~100 ホモ・ハビリス万年前)や、中国(北京原人約35~50 万年 ホモ・サピエンス 前)に拡大した。
ヨーロッパでは、少し遅れて約3万~2.5万年前の地層から、ネアンデルタール人が見つかっている。
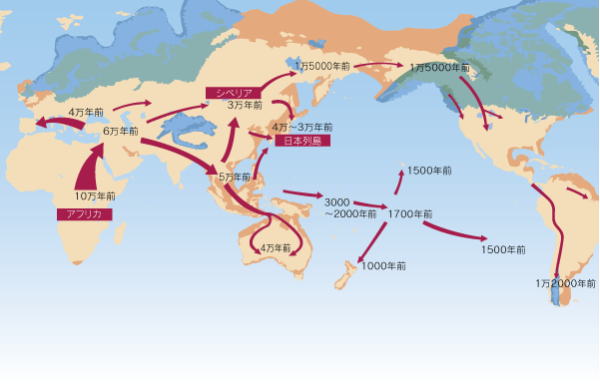
現生人類のホモ・サピエンスは、ミトコンドリアDNA分析の結果から、約20万年前のアフリカで生まれたとされる。ホモ・サピエンスは、厳しい氷期の気候にも適応して、世界各地に生 存領域を広げていった。
ホモ・サピエンスは、約10万年前にアフリカを出て、中東に達し、北のヨーロッパへ向かったグループと、東に向かったグループに分かれた。
東に向かったグループは、南アジアを進み、インドネシアの島嶼伝いにオーストラリアに達し(約5~6万年前)、有袋類のみの世界であったオーストラリアを改変した。
インドから東へ向かったグループは、中国を経由してシベリアには約2.5~3.5万年前に到達し、更にベーリング海峡を渡って、約1万2千年前には北アメリカに到達した。
ホモ・サピエンスは、地上で最強の猛獣であり、多くの動物を狩猟の対象とした。
多くの大型動物が約1万年前に絶滅したが、丁度氷期から間氷期に移行する時期に相当し、気温の変化により植生が変わって、食物等がなくなって絶滅した種もあるが、人類によって滅ぼされた種もあると見られている。
最近数百年間でも、ドードーやステラーカイギュウなどのように、人類によって短期間に狩りつくされた種がある。第四紀の哺乳類全体の傾向として、新第三紀に比べて、種や個体数が減少したことがあげられる。
長鼻目は一時は南極とオーストラリアを除く、全世界に分布したが、現在はインドとアフリカに2種を残すのみ、奇蹄類のサイも、現生種は5種、同じく奇蹄類のウマ類も種数を大幅に減らした。
#目 次
地球7つの秘密 完
|
|


